raymond f. gasser
ATLAS OF HUMAN EMBRYOS
Copyright © 1975 RF Gasser, PhD. All rights reserved. Reproduced with permission.Chapter IV (Pages 24-46)
the fourth week of life
somite and neural tube period

FIG. 4–1
Dorsal aspect of the 10-somite embryo. [printable version of FIG 4-1]
I. EMBRYO PROPER
A. EXTERNAL APPEARANCE
Figs. 4–1, 4–2The specimens measure approximately 1 to 3.5 mm in length and have 1 to 29 pairs of somites.
The head and tail folds move the attachment of the amnion to the ventral side of the head and tail regions, respectively. The lateral body folds move the amnion attachment to the ventrolateral surface in the midportion of the embryo.
The head region is elevated above the yolk sac by the large pericardial sac, the midportion lies upon the yolk sac and the caudal region is curved toward the yolk sac.
The embryo possesses somites, which are apparent through the ectoderm.
The neural tube develops from the neural plate and remains open at each end for 2 to 4 days.
There may be two dorsolateral enlargements in the head region, the trigeminal and otic prominences, separated by the preotic sulcus. The trigeminal prominence is continuous ventrally with the broad first branchial arch that extends to the pericardial sac. The otic prominence is continuous ventrally with an area that will soon become the second branchial arch.
B. ECTODERM
Figs. 4–1, 4–2ANERVOUS SYSTEM
The neural plate is transformed into the neural tube as a result of the neural folds joining in the midline dorsally. The rostral and caudal openings of the tube are called neuropores. The rostral neuropore closes between 18 and 20 somites. The caudal neuropore closes at 25 somites.
Brain
Three brain subdivisions are present in the cranial portion of the tube and are named, from cranial to caudal, the prosencephalon, mesencephalon and rhombencephalon. The boundary between the pros- and mesencephalon is demarcated by a ventral bend, called the cephalic flexure. An external groove and a prominent swelling on the medial surface of the neural plate may also demarcate the boundary. The boundary between the mes- and rhombencephalon is distinguished by a groove on the medial and lateral surfaces of the neural plate or tube.
Prosencephalon
The medial surface possesses a broad, shallow depression, called the optic groove, where the optic cup will form by evagination after closure of the neural tube. This is the first appearance of the eye. The optic chiasmal area is ventral where the two walls come together.
Mesencephalon
The mesencephalon is a short segment of the neural plate or tube which bulges internally and externally. Its dorsal portion will form the tectum; its floor and medial wall will give rise to the tegmentum and basis pedunculi area.
Rhombencephalon
The rhombencephalon extends from the mesencephalon to the level of somite 4 where it is continuous with the spinal cord segment. It can be subdivided into preotic and postotic portions by placode VIII (otic).
Neural Crests
Collections of cells called neural crests stream from the angle between the neural and superficial ectoderm. The following crests can be recognized during the early part of the fourth week.
Trigeminal Crest
The trigeminal crest is the most rostral of the neural crests and is located at the level of the mesencephalon and the preotic part of the rhombencephalon. Initially, it is the largest of all the neural crests, forming most of the trigeminal prominence. It spreads laterally and is gradually lost in the mesenchyme of the first branchial arch. Its caudal part is closely associated with placode V.
Facioacoustic Crest
The facioacoustic crest is a mass at the level of the otic part of the rhombencephalon and constitutes most of the otic prominence. It blends with mesenchyme of the otic region. Its peripheral portion is associated with placode VII and placode VIII (otic).
Glossopharyngeal–Vagal Crest
The glossopharyngeal–vagal crest develops midway between the rostral neuropore and somite 1 at the level of the postotic part of the rhombencephalon. Initially it is not well defined and will later become two separate crests. It is closely associated with the deep surface of placode IX–X.
Occipitospinal Crests
One occipitospinal crest will develop in each body segment between the neural tube and each somite. Initially, it is poorly defined and its cells become scattered among the mesenchymal cells.
ECTODERMAL PLACODES IN THE HEAD REGION
Localized thickenings of the ectoderm called placodes develop on the lateral side of the head in the branchial arch region. They are closely related to the cranial neural crests.
Placode V
Placode V overlies the caudal edge of the trigeminal crest. Its boundaries are poorly defined and it disappears early.
Placode VII
A band of thickened ectoderm represents placode VII and extends from the rostral side of placode VIII (otic) to the first branchial groove. It is also known as the epibranchial placode of the second branchial arch and is associated with the facial part of the facioacoustic crest.
Placode VIII (Otic)
Placode VIII (otic) is the most conspicuous area of ectodermal thickenings and represents the first appearance of the ear. It is located on the summit of the otic prominence and is demarcated caudally by the shallow postotic sulcus. It is associated with the acoustic part of the facioacoustic crest.
Placode IX–X
A tongue shaped area of ectodermal thickening represents placode IX-X and extends caudally from placode VIII to the level of somite 1. Initially, it is associated with the combined glossopharyngeal-vagal crest but soon subdivides into two epibranchial placodes when the IX-X crest becomes distinct and separate.
NOTOCHORD
Fig. 4–2AThe notochord develops from the notochordal process and extends in the midline from the cranial end of the foregut to the tail. Initially it is a plate or ∩ shaped and at most levels becomes the dorsal part of the gut lining. It is in contact with the neural tube dorsally and terminates in the head region near the pros- and mesencephalic junction. In the tail region the notochord separates from the gut endoderm, becomes cylindrical and unites with the underside of the neural plate.

FIG. 4–2 [printable version of FIG 4-2]
- Midsagittal view of the 10-somite embryo showing the relations of the neural tube, notochord, gut and pericardial sac.
- Left lateral view of the 10-somite embryo showing the cardiovascular system. Arrows indicate the position of head and tail folds.
C. ENDODERM
Fig. 4–2AWith the development of the head, tail and lateral body folds, the endoderm that previously formed the roof of the yolk sac is molded into blind tubes in the head and tail regions known as the foregut and hindgut, respectively. The intermediate endoderm that remains as the roof of the yolk sac forms the midgut. These three endodermal subdivisions represent the first appearance of the Alimentary and Respiratory Systems.
FOREGUT
The foregut is compressed in the horizontal plane with its lateral portion extending dorsally. Its cranial end terminates blindly at the oropharyngeal membrane. Caudally it opens into the midgut at the anterior (cranial) intestinal portal.
Ventral outpouchings
-
The buccal pouch is a slight bulge into the oropharyngeal membrane area.
The thyroid diverticulum is the most prominent and deepest outpouching and is located in the midline at the level of the first aortic arches.
A midline trough called the laryngotracheal groove extends caudally from the thyroid diverticulum to the anterior intestinal portal. At its caudal termination the endodermal lining forms a midline bud that represents the unpaired lung primordium (Respiratory System).
Lateral outpouching
-
The first pharyngeal pouch is a clearly defined bulge extending laterally and dorsally around the side of the dorsal aorta. The closing membrane of the first branchial groove is a small area caudal to pouch 1 where ectoderm and endoderm are in contact.
HINDGUT
Caudally the short, blunt hindgut ends blindly at the cloacal membrane. It opens at its cranial end into the midgut at the posterior (caudal) intestinal portal. The allantoic diverticulum (allantois) arises from the ventral side of the hindgut and passes into the connecting stalk.
MIDGUT
The endoderm of the midgut constitutes the roof of the yolk sac cavity between the cranial and caudal intestinal portals. The septum transversum lies ventral to the cranial intestinal portal where the hepatic diverticulum forms.
D. MESODERM
Mesoderm is located on each side of the midline and can be subdivided by its location into four portions: head, paraxial, intermediate and lateral.
HEAD MESODERM
Mesenchymal tissue collects throughout the head region beneath the ectoderm. It probably arises from the cranial neural crests and the primitive streak.
PARAXIAL (SOMITIC) MESODERM
The paraxial mesoderm is located on each side of the neural tube where it collects into blocks of epitheloid cells called somites. The somites are triangular shaped on transverse section with dorsal, ventral and medial walls. The epitheloid cells are arranged around a central cavity called the myocoele.
Somites are segmentally arranged with the first pair forming near the cranial end of the embryo in the future occipital region. Segmentation occurs in a craniocaudal sequence with their formation closely trailing the closure of the neural tube.
The somite number is used to determine the relative age of embryos in the somite period. Eventually there are 42 to 44 somite pairs formed (4 occipital, 8 cervical, 12 thoracic, 5 lumbar, 5 sacral and 8 to 10 coccygeal).
By the time the more caudal somites appear, the cranial ones break down as a result of proliferation. The somites will give rise to major components of the Skeletal, Muscular and Integumentary Systems.
INTERMEDIATE MESODERM
The intermediate mesoderm temporarily connects the paraxial with the lateral plate mesoderm.
In the cervical segments the intermediate mesoderm is arranged into a group of cells that produce a nonfunctional, vestigial kidney of lower forms called the pronephros. The pronephros represents the first appearance of the Urogenital System and is a very transitory structure.
During the early part of the fourth week, the pronephros is composed of small, isolated groups of cells called pronephric rudiments at the level of somites 3 to 8. At the level of somites 8 to 10 (C-4 to C-6) the cells form a pronephric ridge. There is no definite evidence of segmentation or lumen formation.

FIG. 4–3
Ventral aspect of the heart and pericardial sac of the 10-somite embryo. The aortic sac lies just dorsal to the pericardial cavity. [printable version of FIG 4-3]
LATERAL (PLATE) MESODERM
Lateral mesoderm lies between the intermediate mesoderm and the extraembryonic mesoblasts at the edge of the embryo.
Intercellular clefts appear in the lateral mesoderm and separate it into an outer layer called somatic mesoderm and an inner layer known as splanchnic mesoderm. As the clefts enlarge and coalesce they produce a cavity between the layers on each side known as the embryonic coelom.
In the midportion of the embryo, the splanchnic mesoderm is continuous laterally with the extraembryonic mesoblasts of the yolk sac. The embryonic coelom communicates laterally with the extraembryonic coelom.
The somatic mesoderm becomes closely associated with the overlying ectoderm and together they form a double layer called the somatopleure. The splanchnic mesoderm is similarly associated with the underlying endoderm and together they form the splanchnopleure. The intervening embryonic coelom is continuous cranially through the pericardioperi-toneal canal (coelomic duct) with the pericardial cavity.
HEART
Figs. 4–2, 4–3The heart begins as an endocardial tube that forms in front of the prochordal plate from bilateral angiogenic cell clusters located between the cardiogenic plate and the underlying yolk sac endoderm. The cardiogenic plate is a thickened layer of splanchnic mesoderm that will give rise to the epimyocardium (see below). With the formation of the head fold and the expansion of the prosencephalon, the cardiogenic plate assumes a position ventral to the foregut. The angiogenic cell clusters become a pair of endothelial tubes in the splanchnic mesodermal layer and bulge into the pericardial part of the embryonic coelom. With formation of the lateral body folds, the paired endothelial tubes fuse in the midline producing a single endocardial tube ventral to the foregut.
With continued growth the heart tube assumes an S shape and is suspended from the dorsal wall of the pericardial cavity by a short mesentery called the mesocardium. It is separated from the yolk sac by the septum transversum and is composed of two layers, a thick outer layer called the primitive epimyocardium and a thin inner layer, the primitive endocardium. The two layers are separated by a distinct endomyocardial space, which is filled with cardiac jelly.
The heart tube begins to beat, circulating primitive blood cells through the vessels of the embryo, chorion, connecting stalk and yolk sac. Cranially it is continuous with arteries; caudally it receives the veins.
Three areas can be recognized in the single tube. From cranial to caudal they are as follows.
-
The aortic sac or roughly ovoid, cranial end of the tube from which the aortic arches arise. It lies mainly in the dorsoventral plane outside the pericardial cavity.
The primitive bulbus cordis or ventral portion curving to the right of the midline is separated from the aortic sac by a slight constriction.
The primitive ventricle is the most prominent dilatation, extending to the left and dorsally at right angles to the primitive bulbus cordis from which it is partially separated by the bulboventricular sulcus.
The single tube bifurcates near the caudal end of the pericardial sac into the two, slightly dilated primitive atria that are in the floor of the cranial intestinal portal. The atria are separated from the ventricle by shallow right and deep left atrioventricular (A-V) sulci. Internally, the single ventricular chamber communicates with both atrial chambers through a single atrioventricular canal.
Each atrium is continuous with a dilated chamber on each side of the cranial intestinal portal called the sinus venosus. Each sinus venosus receives veins from the connecting stalk, chorion and yolk sac.
ARTERIES
Fig. 4–2BThe first pair of aortic arches connects the aortic sac with the paired dorsal aortas located on each side, dorsal to the primitive gut. The second pair forms caudal to the first pharyngeal pouch as small, irregular vessels. Cranial to the first arch the dorsal aorta on each side extends toward the pros- and mesencephalon as the internal carotid arteries.
Small sprouts arise from the dorsal aortas along the rhom-bencephalon and between most of the somites. They represent the beginning of dorsal intersegmental arteries.
Each of the dorsal aortas terminates caudally by dividing into many ventral branches that break up into plexuses on the midgut, hindgut and yolk sac. The plexuses connect with the single, large umbilical artery in the connecting stalk that courses to the chorionic villi. The connections on each side will form the right and left umbilical arteries.
VEINS
Fig. 4–2BTributaries of the large vitelline plexus on the yolk sac come together to form right and left vitelline veins that drain into the sinus venosus on the corresponding side.
Blood from the chorion courses through large spaces in the connecting stalk that join together on each side to form right and left umbilical veins. Each vein passes along the lateral side of the embryo where the amnion attaches and empties into the sinus venosus on that side.
Chains of relatively large capillary blood vessels begin forming the primary head vein on the lateral surface of the neural tube in the head region. These channels will eventually become a part of the cardinal system of veins that drain the embryo.
II. EXTRAEMBRYONIC MEMBRANES
Figs. 4–1, 4–2A. YOLK SAC
The entrance from the yolk sac into the primitive gut is ovoid in shape because the umbilical veins constrict the entrance laterally. The yolk sac is covered with a vascular plexus and blood islands that drain to the sinus venosus of the heart by the large umbilical and vitelline veins on each side. The primordial germ cells are located in the wall of the yolk sac near the allantois. They will later migrate in the hindgut mesentery to the dorsal body wall.
B. AMNION
As the head and lateral body folds form, the cranial attachment of the amnion moves to the ventral surface of the embryo where the pericardial cavity meets the yolk sac. As the tail and lateral body folds form, the caudal attachment of the amnion moves to the ventral side where the connecting stalk attaches.
C. CONNECTING STALK
The connecting stalk lies between the amnion and yolk sac when the body folds develop. It contains a large, common umbilical artery and two umbilical veins that conduct blood to and from the chorion.
D. ALLANTOIS
As the tail fold develops, the attachment of the allantois moves ventrally to form part of the ventral wall of the hindgut. It remains a diverticulum lined with endoderm and ends blindly in the connecting stalk.
THE 10-SOMITE EMBRYO
STAGE 10
AGE 22 DAYS
GREATEST LENGTH 3.0 MM
Carnegie collection 8004
Reference
Corner GW: A well-preserved human embryo of 10 somites. Contrib Embryol Carnegie Instn 20:81–99, 1929
Heuser CH, Corner GW: Developmental horizons in human embryos. Description of age group X, 4 to 12 somites. Contrib Embryol Carnegie Instn 36:29–39, 1959
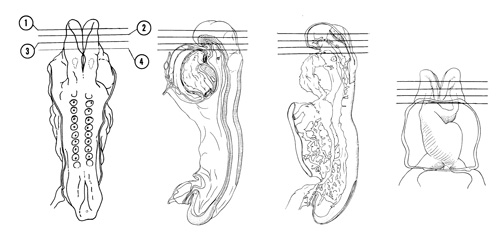
FIG. 4–4
SECTION 1
A section through the open curved part of the prosencephalon showing its junction with the mesencephalon.
Observe:
- The optic area that will form the optic vesicle, which will eventually become the retina of the eye.
- The continuity of the ectoderm with the neural plate.
- The trigeminal neural crest that blends with mesodermal cells in the area.
A section through the ventral part of the prosencephalon at its junction with the mesencephalon.
Observe:
- The primordial area of the optic chiasma in the ventral part of the prosencephalon.
- The middle of the trigeminal crest.
- The large carotid arteries on each side of the brain filled with primitive, nucleated blood cells.
A section through the cranial end of the foregut showing its separation from the exterior by the two-layered, oropharyngeal membrane.
Observe:
- The preotic part of the rhombencephalon that is open dorsally showing the neural plate, groove and folds.
- The caudal part of the trigeminal crest extending laterally toward a slightly thickened area of ectoderm (placode V).
- The cranial end of the foregut lined with endoderm.
- The cranial ends of the dorsal aortas.
- The cranial end of the notochord in the roof of the foregut.
A section through the first branchial arch and the cranial part of the pericardial cavity.
Observe:
- Because of the head fold (see Figs. 4–1 and 4–2B), the attachment of the amnion moves to the ventral side of the head region.
- The cranial edge of the primitive bulbus cordis surrounded by the pericardium on the ventral aspect of the head.
- Within the first branchial arch is the first aortic arch about to join the dorsal aorta.
- The slight ventral bulge in the floor of the foregut forming the buccal pouch.
- The first appearance of the primary head vein between the brain and dorsal aorta.
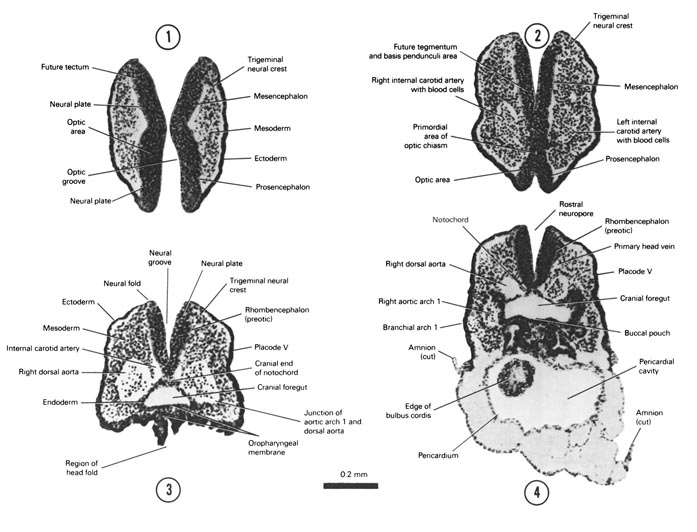
FIG. 4–4 [printable version of FIG 4-4]
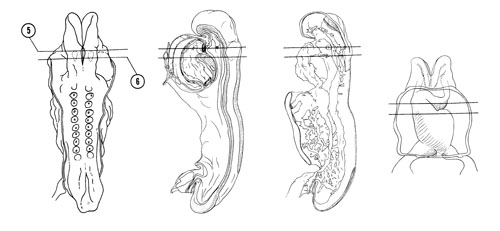
FIG. 4–5
SECTION 5
A section through the thyroid diverticulum and primitive bulbus cordis that is to the right of the midline.
Observe:
- The close relation of the thyroid diverticulum to the aortic sac and first aortic arch.
- The first pharyngeal pouch with its dorsal extension.
- The closing membrane of the first branchial groove where the tympanic membrane (eardrum) will develop.
- The facioacoustic crest with cells extending toward placode VII.
- The abnormally distended pericardial cavity.
A section through placode VIII (otic) and cranial part of the primitive ventricle of the heart.
Observe:
- The two layers of the heart, primitive endocardium and epimyocardium, separated by the endomyocardial space.
- The heart tube suspended by the mesocardium.
- The laryngotracheal groove in the floor of the foregut.
- The close relation between the caudal part of the facioacoustic crest and placode VIII (otic), which will form the membranous labyrinth of the internal ear.
- The caudal extent of the rostral neuropore.
 and cranial part of the primitive ventricle of the heart.")
FIG. 4–5 [printable version of FIG 4-5]
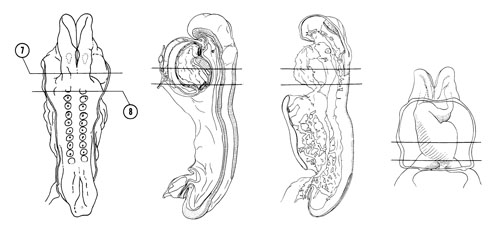
FIG. 4–6
SECTION 7
A section through the middle of the primitive ventricle of the heart that is to the left of the midline.
Observe:
- The close relation between the mesocardium and the laryngotracheal groove in the floor of the foregut.
- The close relation between the glossopharyngeal-vagal neural crest and placode IX–X.
- The closed neural tube at the postotic part of the rhombencephalon.
A section through somite 1, the primitive atria of the heart and the cranial edge of the yolk sac cavity.
Observe:
- The separation of the pericardial cavity from the yolk sac cavity by the septum transversum.
- The narrow atrioventricular canal that connects the two atria to the primitive ventricle in Section 7.
- The close relation of foregut endoderm to the primitive atria.
- The close association of the neural tube, somite 1 and the dorsal aorta.
- The attachment of the amnion to the lateral aspect of the pericardium.

FIG. 4–6 [printable version of FIG 4-6]
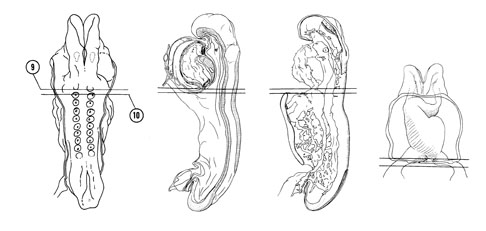
FIG. 4–7
SECTION 9
A section through the caudal part of somite 1 and the pericardial cavity at the level of the lung primordium.
Observe:
- The reduction in the size of the endomyocardial space.
- The cranial edge of the left sinus venosus where it becomes continuous with the primitive left atrium.
- The close relation of the unpaired lung primordium to the heart.
- The division of mesoderm into paraxial (somitic), intermediate and lateral portions.
- The subdivision of lateral mesoderm into splanchnic and somatic portions.
A section through somite 2, the sinus venosus and the fore- and midgut junction.
Observe:
- The anterior intestinal portal at the junction of the foregut and midgut.
- The middle of the septum transversum where the hepatic diverticulum is about to form.
- The sinoatrial orifice.
- The pericardioperitoneal canal on each side of the gut which connects the pericardial cavity in Section 9 with the coelomic cavity in Figure 4–8, Section 14.
- A few neural crest cells in the future occipital region.

FIG. 4–7 [printable version of FIG 4-7]
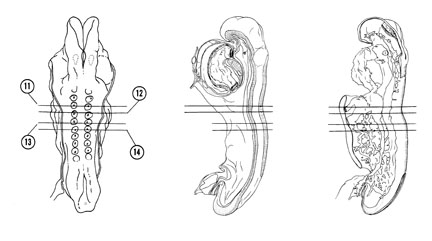
FIG. 4–8
SECTION 11
A section through somite 3 and the cranial part of the midgut.
Observe:
- The wide communication between the midgut and yolk sac cavity.
- The junction of the umbilical and vitelline veins in Section 12 with the sinus venosus.
- The pericardioperitoneal canal surrounded with lateral plate mesoderm.
- The ventromedial extent of the somites below the neural tube.
A section through somite 4.
Observe:
- The vitelline veins on the lateral wall of the yolk sac.
- The large umbilical veins lateral to the pericardioperitoneal canal.
- A pronephric rudiment between the coelomic duct and the somite.
- The close association between the neural crest cells and the cells of the somite.
A section through somite 5.
Observe:
- The midgut endoderm forming the roof of the yolk sac cavity as in earlier stages.
- The near junction of the pericardioperitoneal canal with the extraembryonic coelom.
- The spinal cord part of the neural tube.
A section through the middle of somite 6.
Observe:
- The notochord changing from a plate in the roof of the gut to a rounded shape.
- The large myocoele in the center of the somite filled with a clump of cells.

FIG. 4–8 [printable version of FIG 4-8]
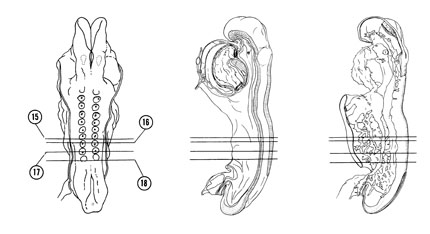
FIG. 4–9
SECTION 15
A section through somite 7.
Observe:
- The lateral body fold that narrows the communication between the midgut and the yolk sac cavity.
- The dorsal aorta adjacent to the roof of the midgut on each side.
- The neural canal that will become the central canal of the spinal cord.
A section through somite 8.
Observe:
- Part of the vitelline plexus in the wall of the yolk sac cavity.
- The extraembryonic coelom lined with extraembryonic mesoblasts.
- The coelomic cavity subdividing the lateral plate mesoderm.
- The pronephros represented by a ridge in the intermediate mesoderm.
A section through somite 9 and the caudal part of the midgut.
Observe:
- The communication between the coelomic cavity and extraembryonic coelom.
- The three subdivisions of mesoderm between the ectoderm and endoderm.
- The separation of lateral plate mesoderm into splanchnic and somatic parts by the embryonic coelom.
A section through somite 10 and the junction of midgut with hindgut.
Observe:
- The posterior intestinal portal where the gut no longer opens ventrally into the yolk sac cavity.
- The umbilical veins along the lateral borders of the embryo.
- The more lateral position of the dorsal aortas.
- The pronephric ridge in the intermediate portion of the mesoderm.

FIG. 4–9 [printable version of FIG 4-9]
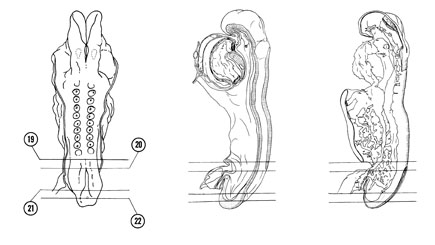
FIG. 4–10
SECTION 19
A section through the caudal part of the neural tube.
Observe:
- The cranial part of the hindgut.
- The splanchnopleure formed from gut endoderm and splanchnic mesoderm.
- The somatopleure formed from ectoderm and somatic mesoderm.
- The embryonic coelom separating splanchnopleure and somatopleure.
A section through the connecting stalk and the junction of the allantois with the floor of the hindgut.
Observe:
- The junction of the dorsal aortas with the vitelline plexus and the junction of the plexus with the common umbilical artery.
- The common umbilical artery in the connecting stalk and the left and right umbilical veins leaving the stalk.
- The attachment of the amnion to the stalk.
- The caudal extent of the coelomic cavity.
- The caudal neuropore where the neural plate has not yet closed.
A section through the embryo caudal to the tail fold at the level of the cloacal membrane.
Observe:
- The embryo completely clothed in ectoderm.
- The fusion of endoderm and ectoderm at the cloacal membrane in the floor of the hindgut.
- The thick, solid layer of mesoderm.
- The caudal end of the notochord where it has a rounded appearance on cross section.
- The neural folds and groove.
A section through the caudal tip of the embryo.
Observe:
- The primitive streak where mesodermal cells are being formed.
- The neuroectodermal junction where the neural plate is in contact with ectoderm.
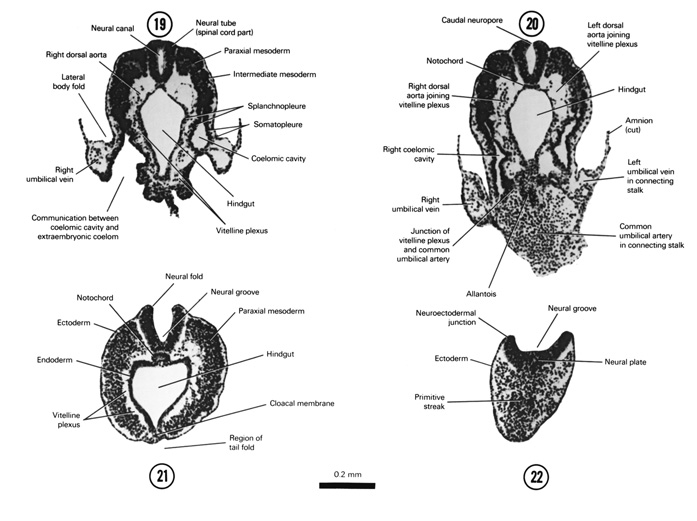
FIG. 4–10 [printable version of FIG 4-10]