raymond f. gasser
ATLAS OF HUMAN EMBRYOS
Copyright © 1975 RF Gasser, PhD. All rights reserved. Reproduced with permission.Chapter VI (Pages 103-162)
- External Appearance
- Nervous System
- Alimentary and Respiratory Systems
- Urogenital System
- Coelomic Cavity
- Mesenteries
- Cardiovascular System
- Skeletal System
- Muscular System
- Integumentary System
- Special Sense Organs
the sixth week of life
late branchial arch and limb bud period
I. EXTERNAL APPEARANCE
Fig. 6–1AThe specimens measure approximately 8 to 14 mm in length.
The head region greatly increases in size. The second branchial arch grows caudally forming the opercular fold that partially covers the third and fourth branchial arches in the floor of the cervical sinus. The optic cup and lens vesicle, as well as various areas of the brain, can be located on the surface. The olfactory placode sinks below the surface in the floor of the nasal pit. The surface features of the face are described in Section III, A.
The limb buds become finlike with a constriction around the middle portion, which separates off a distal segment called the primitive hand or foot plate.
The heart and liver form prominences in the ventral wall between the branchial arches and umbilical cord.
The amnion fuses with the surface of the large umbilical cord. Its attachment moves to the distal part of the cord and is no longer evident on the ventral body wall. The yolk stalk becomes incorporated into the cord and usually disappears.
The surface features of the external genitalia (perineal region) between the lower limb and tail buds are described in Section IV, E.
The tail bud becomes more pointed and begins to disappear.
II. NERVOUS SYSTEM
A. CENTRAL NERVOUS SYSTEM (CNS)
Fig. 6–1BGENERAL
The cephalic and cervical flexures become more pronounced as a sharp dorsal bend called the pontine flexure appears in the middle of the rhombencephalon.
The brain and spinal cord are surrounded by a loose network of cells called the primitive meninx from which develop the meninges (protective membranes) and some bones of the skull.
BRAIN
Telencephalon
A cerebral vesicle forms on each side as an outpouching of the lateral wall leaving a midportion called the telencephalon medium.
Cerebral Vesicle
Each vesicle can be subdivided into a thick basal striatal part and a thin dorsal suprastriatal part. The rhinencephalon or smell brain that is composed of several regions of the telencephalon is becoming apparent. The primordial tuberculum olfactorium is the clear, marginal area on the ventral surface of the cerebral vesicle. In front of this area the olfactory fila from the olfactory epithelium are about to enter the telencephalon. The primordial hippocampal formation is represented by a) a clear, marginal layer in the dorsal part of the cerebral vesicle called the primordial gyrus dentatus and b) the more dense area immediately lateral called the primordial cornu Ammonis.Telencephalon Medium
The floor of the telencephalon medium is composed of the thin lamina terminalis and the preoptic area, both of which are located just rostral to the chiasmal area of the diencephalon. The roof of the telencephalon medium is continuous with the roof of the diencephalon and with it constitutes the roof of the third ventricle. Near the junction with the diencephalon the roof bulges to produce the paraphyseal arch.Prosocoele
That part of the prosocoele that extends into the cerebral vesicle on each side will become the lateral ventricle. The part remaining in the midline becomes the third ventricle. The interventricular foramen is developing in the junction region.
Diencephalon
The sulcus dorsalis appears in the lateral wall and separates the dorsal thalamus from the epithalamus. The roof of the epithalamus evaginates to produce the pineal bud.
The sulcus medius and sulcus ventralis become better defined. The sulcus ventralis continues rostrally as the optic groove, which passes into the optic stalk.
Two ventral bulges develop in the hypothalamus: a) a rostral, midline bulge, the neurohypophyseal bud, adjacent to the hypophyseal pouch and b) a caudal bulge on each side called the mammillary area. The third ventricle above the mammillary area forms the mammillary recess. The marginal area increases in thickness in the optic chiasmal area.
Mesencephalon
The sulcus limitans is very prominent and divides the lateral wall into a dorsal alar plate and a ventral basal plate.
Three adult subdivisions—The alar plate on each side together with the roof plate form the tectum. The basal plate on each side together with the floor plate form the tegmentum. The marginal layer ventral to the tegmentum increases in width producing the basis pedunculi area (crus cerebri).
The caudal, constricted portion that is continuous with the rhombencephalon is called the isthmus.
The mesocoele connects the third ventricle with the rhombocoele.
Rhombencephalon
The rhombocoele dilates into a diamond shaped chamber forming the fourth ventricle. It is continuous caudally with the narrow neural canal of the spinal cord. The thin roof of the fourth ventricle also enlarges and becomes diamond shaped.
The basal plates are separated in the midline by a deep median sulcus. The alar plates become separated as the roof enlarges and move to a position dorsolateral to the basal plates. The sulcus limitans marks the boundary between the alar and basal plates.
Rhombomeres are sometimes apparent on the inner surface of the ventral wall but disappear on the outer surface.
The pontine flexure divides the rhombencephalon into a rostral portion, the metencephalon, and a caudal portion, the myelencephalon.
Metencephalon—The marginal layer of the basal plate begins to thicken to produce the pons area. The lateral edge of the alar plate is called the rhombic lip where the cerebellum will later develop.
Myelencephalon—The lateral wall of the myelencephalon is divided into large alar and basal plates. In the rostral part of the myelencephalon the alar plates are separated from each other by the roof of the fourth ventricle. Here they are in a position dorsolateral to the basal plates and contribute to the formation of the floor of the fourth ventricle. In the caudal part of the myelencephalon the alar plates approach each other and are separated only by a narrow strip of roof plate. The marginal layer of the basal plate begins to thicken to form the pyramidal tract area.

FIG. 6–1 [printable version of FIG 6-1]
- External features of the 10-mm embryo.
- Central nervous system of the 10-mm embryo. Arrows indicate the position of the cephalic, pontine and cervical flexures.
SPINAL CORD
The spinal cord enlarges and extends into the tail bud where it terminates adjacent to the ectoderm dorsally. The most caudal segment represents the primitive filum terminale, which will increase in length when the spinal cord fails to keep pace with the longitudinal growth of the vertebral column.
The ependymal, mantle and marginal layers become very distinct. The neuroblasts are mainly in the mantle layer. The basal portion of the mantle layer is thick and has a marginal layer of uniform thickness at its periphery. The marginal layer is thickest dorsally where the dorsal roots from spinal ganglia enter the cord. The floor plate thickens but the roof plate remains thin.
B. PERIPHERAL NERVOUS SYSTEM
Fig. 6–2BCRANIAL NERVES
All twelve cranial nerves can be identified.
The olfactory nerve (I) is represented as fila among a cellular condensation between the olfactory epithelium and the ventral surface of the cerebral vesicle.
The optic nerve (II) is in the form of the optic stalk that joins the optic cup to the diencephalon. The stalk lumen begins to narrow.
The oculomotor (III), trochlear (IV) and abducens (VI) nerves appear as small fascicles that terminate in premuscle condensations dorsal to the optic cup.
-
Each of the four cranial neural crests transforms into distinct cranial nerves with motor and sensory components. The sensory ganglion (or ganglia) on each nerve becomes distinct and loses contact with its epibranchial placode. The placode disappears as the ganglion forms.
The three major divisions or branches of the trigeminal nerve (V) become apparent. The ophthalmic nerve (V1) passes dorsal to the optic stalk, the maxillary nerve (V2) courses into the maxillary process and the mandibular nerve (V3) proceeds into the mandibular process.
The facial part of the facioacoustic neural crest gives rise to the facial nerve (VII). The greater petrosal and chorda tympani branches can be traced into the first branchial arch where they communicate with the maxillary and mandibular nerves, respectively. The acoustic part of the facioacoustic neural crest contributes to the formation of the vestibulocochlear nerve (VIII). The vestibular and cochlear (spiral) ganglia develop in the region where the crest cells make contact with the otocyst.
The glossopharyngeal nerve (IX) passes through the third branchial arch and terminates in the floor of the pharynx just caudal to the foramen cecum linguae.
The vagus nerve (X) gives rise to its superior laryngeal and recurrent (inferior) laryngeal branches. It can be traced as far caudally as the stomach. The cranial part of the accessory nerve (XI) becomes a part of the vagus nerve and its recurrent laryngeal branch.
The spinal accessory nerve (XI) becomes a very distinct fascicle, which terminates in the sternocleidomastoid-trapezius premuscle mass dorsal to the cervical sinus.
The hypoglossal nerve (XII) appears as a large, discrete fascicle passing lateral to the vagus nerve and terminating in the premuscle masses of the tongue. It forms communications with the upper three cervical spinal nerves.
SPINAL NERVES
Sensory neuroblasts differentiate from neural crest cells and aggregate in each segment to form spinal ganglia. Each neuroblast initially develops two processes, one called the axon, which grows centrally and penetrates the dorsal surface of the spinal cord, and another called the dendrite, which grows into the periphery. The two processes later will join together near the cell body of the neuroblast and become one process (unipolar). The axons from each ganglion collect together to form the dorsal (sensory) root of the spinal nerve. The dendrite reaches the periphery by passing through the spinal nerve and its branches. Sensory receptors later will develop on many of the dendritic terminals.
Motor neuroblasts differentiate from cells in the basal plate portion of the mantle layer. They will form multiple processes (multipolar), one axon but many dendrites. The axon grows through the marginal layer to emerge from the spinal cord. Axons in each segment collect together to form the ventral (motor) root of the spinal nerve. They course to the periphery through the spinal nerve and its branches along with the dendrites of the sensory neuroblasts. They terminate on or near skeletal premuscle cells.

FIG. 6–2 [printable version of FIG 6-2]
- Peripheral nervous system of the 10-mm embryo.
- Alimentary and respiratory systems of the 10-mm embryo.
The definitive number of spinal ganglia (31 pairs) becomes evident. The dorsal and ventral primary rami of the spinal nerves can be identified. The dorsal primary rami will innervate the deep muscles of the back and the overlying subcutaneous tissue and dermis. The ventral primary rami will innervate the muscles, subcutaneous tissue and dermis of the limbs and ventrolateral body wall.
The ventral primary rami join together in all except the thoracic segments to form plexuses where nerve processes intermingle. They behave as indicated in Table 6–1.
| Table 6–1. Nerve Plexuses | ||
| Segments | Plexus | Region Innervated |
|---|---|---|
| C-1, C-2, C-3, C-4 | Cervical | Ventrolateral neck |
| C-5, C-6, C-7, C-8, T-1 | Brachial | Upper limb and shoulder |
| T-1 through T-12 | Eleven intercostal nerves and one subcostal nerve (T-12) (no plexus formed) | Ventrolateral thoracic and abdominal walls |
| L-1, L-2, L-3, L-4 | Lumbar | Lower limb and groin |
| L-4, L-5, S-1, S-2, S-3 | Sacral | Lower limb and perineum |
| S-4, S-5, Coccygeal | Coccygeal | Coccygeal (tail) region |
The phrenic nerve (C-3, C-4, C-5) becomes a discrete nerve fascicle, which passes lateral to the heart and terminates in the septum transversum where the diaphragm develops.
C. AUTONOMIC NERVOUS SYSTEM
Fig. 6–2AFUNDAMENTALS
The nerves of the autonomic nervous system course with the peripheral nervous system and consist of all the motor neurons that innervate visceral structures (viz., glands and smooth and cardiac muscles).
A unique feature of this system is that the messages from the CNS must travel through not one but two motor neurons before they reach the peripheral effector organ. The cell body of the first neuron is located inside the CNS and is called the preganglionic neuron. Its axon leaves the CNS and synapses with the second motor neuron called the postganglionic neuron. Postganglionic neurons are located outside the CNS and collect together to form motor (autonomic) ganglia. The synapse of the preganglionic neuron on the postganglionic neuron occurs in these ganglia.
The autonomic nervous system is divided into two parts depending upon the location of the cell body of the preganglionic neuron within the CNS. If the neuron cell body resides in the thoracic or upper lumbar segments of the spinal cord, it belongs to the sympathetic system; if the neuron cell body resides in either the brain or sacral segments of the spinal cord, it belongs to the parasympathetic system.
SYMPATHETIC NERVOUS SYSTEM
These preganglionic neurons differentiate in the mantle layer of the lateralmost part of the basal plate in the thoracic and upper lumbar segments of the spinal cord (T-1 to L-2). Their axonal processes grow out of the spinal cord through the ventral roots, then enter the spinal nerve. They leave the spinal nerves through the communicating rami and make contact with the postganglionic neurons located in the vicinity of the dorsal aorta.
Neural crest cells give rise to the postganglionic neurons, many of which migrate ventrally to collect into a longitudinal column on each side of the dorsal aorta to form the sympathetic trunk. Some of the neuroblasts migrate beyond the trunk and collect in front of the dorsal aorta forming preaortic ganglia. The axonal process of the postganglionic neurons terminates in viscera.
The suprarenal gland is closely related to the sympathetic nervous system and develops in the upper thoracic segments. It is subdivided into a cortex and a medulla, each having a different origin. The cortex forms from mesothelial cells at the base of the dorsal mesogastrium. The medulla develops from cells that are closely related to sympathetic neuroblasts. They are referred to as prechromaffin cells because they will stain later with chrome salts. These cells differentiate from neural crest cells that collect dorsomedial to the cortical cells and are closely related to similar cells surrounding the celiac branch of the dorsal aorta.
The functions of the sympathetic system are associated with fright and flight reactions.
PARASYMPATHETIC NERVOUS SYSTEM
These preganglionic neurons differentiate in the mantle layer in the lateral part of the basal plate in either the brain or sacral segments of the spinal cord. The axonal process of those neurons in the brain leave by way of cranial nerves III, VII, IX or X. Those in the sacral segments of the spinal cord leave by way of the S-2, S-3 or S-4 ventral roots and travel to the viscera by way of the pelvic splanchnic nerves. The axonal processes synapse on postganglionic neurons close to or within the viscera.
Postganglionic neurons probably arise from neural crest cells in the brain and sacral regions and migrate close to or within the viscera they will innervate. Their axonal processes are short.
The functions of the parasympathetic system are generally antagonistic to those of the sympathetic system.
III. ALIMENTARY AND RESPIRATORY SYSTEMS
Fig. 6–2BA. ORONASOFACIAL REGION
The olfactory placode sinks below the surface to form the floor of a depression called the nasal (olfactory) pit. Proliferating mesenchymal cells around the pit raise the surrounding ectoderm into medial and lateral nasal elevations. The olfactory placode becomes the olfactory epithelium under which mesenchymal cells collect to form the nasal capsule. The frontonasal area lies between as well as dorsal to the medial nasal elevations.
The maxillonasal (nasolacrimal) groove appears in the ectoderm between the lateral nasal elevation and the maxillary process. It extends from the medial side of the optic cup to the nasal pit.
The maxillary process is separated from the medial nasal elevation by an epithelial plate called the nasal fin.
The distal end of the hypophyseal pouch remains in close association with the neurohypophyseal bud of the diencephalon just caudal to the chiasmal area. The pouch elongates and flattens in the transverse plane. Its wall thickens and remains in continuity with the epithelial lining of the stomodeum. The lumen of the pouch communicates with the stomodeum through a narrow constriction that is closing.
B. FOREGUT
The pharyngeal pouches remain connected to the primitive pharynx. The first and second pouches dilate in close proximity to their respective branchial groove. The lining of the third and fourth pouches thickens, becomes removed from the grooves and divides into dorsal and ventral portions. The fifth pouch (ultimobranchial body) is incorporated into the fourth pouch.
The caudal segment of the thyroid diverticulum becomes a solid mass of cells that forms the thyroid gland. It is temporarily connected to the floor of the pharynx at the foramen cecum linguae by the thyroglossal duct.
Swellings appear in the floor of the primitive pharynx where the tongue develops. There are three such swellings rostral to the foramen cecum linguae at the level of the first branchial arch, a midline tuberculum impar flanked on each side by a mandibular swelling. Three similar swellings also form caudal to the foramen cecum linguae at the level of the second, third and fourth branchial arches, a midline hypobranchial eminence flanked on each side by a hyoid swelling.
Mesenchymal cells begin to form a condensed layer peripheral to the pharyngeal endodermal lining.
At the level of the septum transversum the esophagus passes to the left of the midline to terminate in the stomach. The endoderm of the stomach expands into a crescent shape with greater and lesser curvatures surrounded by a dense cellular condensation. The body and pyloric portions of the stomach become evident as rotation begins. The greater curvature (dorsal border) moves to the left and the lesser curvature (ventral border) moves to the right. The dorsal mesogastrium also swings to the left since it is attached to the greater curvature. Mesenchymal cells collect in its caudal portion forming a bulge that represents the primordium of the spleen.
The most caudal segment of the foregut does not dilate like the stomach segment and represents the cranial part of the duodenum. The dorsal pancreatic bud that arises from this segment branches as it invades the caudal part of the dorsal mesogastrium forming the dorsal pancreas. As the bud grows toward the left, it passes ventral to the superior mesenteric vein and terminates close to the splenic primordium.
RESPIRATORY SYSTEM
A third midline swelling appears caudal to the hypobranchial eminence at the level of the fourth branchial arch and represents the epiglottis. The laryngeal orifice caudal to the epiglottis is flanked on each side by arytenoid swellings. A cellular condensation surrounds the primitive larynx and the tracheal endodermal tube.
The trachea grows caudally and bifurcates in the caudal cervical segments into right and left primary bronchi. The right bronchus subdivides into upper, middle and lower lobar buds (secondary bronchi); the left bronchus subdivides into upper and lower lobar buds.
C. MIDGUT
The distal part of the hepatic diverticulum transforms into a complex of branching cords of cells (trabeculae) separated by venous channels (sinusoids), which together make up the liver. The liver lies in the most cranial part of the abdominal cavity and remains separated from the heart by the septum transversum. The proximal part of the hepatic diverticulum becomes the bile duct that empties into the fore- and midgut junction. The gall bladder forms as a diverticulum of the bile duct on the caudal surface of the liver. The bile duct then continues to the liver as the hepatic duct. The ventral pancreas is represented as a branching duct that moves first to the right side of the duodenum then to a dorsal position. It grows into the dorsal mesoduodenum and lies to the right of the portal vein. The ventral pancreatic duct joins the bile duct near the duodenum.
The yolk stalk disappears completely, separating the midgut from the yolk sac.
The midgut increases in length at such a rapid rate that it extends as a loop into the proximal part of the umbilical cord. The superior mesenteric artery courses through the dorsal mesentery to supply all of the midgut derivatives. The midpoint of the loop can be used to divide it into cranial and caudal limbs. The loop rotates counterclockwise around the artery with the cranial limb moving to the right and the caudal limb to the left.
A swelling forms in the caudal limb of the midgut that represents the primitive cecum.
The midgut endodermal tube is surrounded by a dense layer of mesenchymal cells.
D. HINDGUT
The cranial part of the hindgut remains in the midline suspended from the dorsal body wall by a mesentery through which courses the inferior mesenteric artery. The caudal end of the hindgut becomes the rectum and the urogenital sinus, which are separated by the urorectal septum. All of the hindgut derivatives with the exception of the urogenital sinus will be supplied by the inferior mesenteric artery.
The cloacal membrane becomes a vertical plate of epithelium separating the lumen of the cloaca, urogenital sinus and rectum from the exterior.
The tailgut disappears.
The hindgut endodermal tube becomes surrounded by a dense layer of mesenchymal cells.
IV. UROGENITAL SYSTEM
Fig. 6–3A. MESONEPHROS
Many glomeruli and excretory tubules differentiate in the mesonephros causing it to project as a ridge from the dorsal body wall in the C-7 to L-3 segments.
The mesonephros and the more medial gonad are attached to the dorsal body wall by a short, thick urogenital mesentery.
The excretory tubules drain into an enlarged mesonephric duct coursing caudally on the lateral surface of the mesonephros.
The mesonephric duct on each side terminates in its respective horn of the urogenital sinus.
B. METANEPHROS
A minute outpouching called the metanephric diverticulum (ureteric bud) develops on the urogenital sinus horn and grows dorsocranially. Its blind termination dilates to form the primitive renal pelvis, which is surrounded by a cellular condensation called the metanephrogenic mass (blastema).
The metanephrogenic mass is at the level of the L-3 and L-4 segments.

FIG. 6–3
Urogenital system of the 10-mm embryo. [printable version of FIG 6-3]
C. GONAD
The gonadal ridge is very prominent medial to the mesonephros in the T-3 to L-2 segments.
Cells begin to condense adjacent to the surface epithelium
D. UROGENITAL SINUS
As the urogenital sinus is separated from the rectum, it dilates and becomes a large chamber with two horns, each of which is continuous with its respective mesonephric duct.
A constriction develops at the junction of the sinus with the allantois and gradually obliterates the lumen, thus producing an apex in the sinus.
E. EXTERNAL GENITALIA
A midline mesenchymal condensation raises the ectoderm into the genital tubercle that lies between the umbilical cord and the cloacal membrane.
A midline urogenital groove extends caudally from the tubercle and is flanked on each side by a urogenital fold. The cloacal membrane in the floor of the groove separates the cloaca from the exterior. Lateral to the urogenital fold is another mesenchymal condensation, the labioscrotal swelling.
V. COELOMIC CAVITY
A. PERITONEAL CAVITY (ABDOMINAL PORTION)
The communication between the peritoneal cavity and the extraembryonic coelom disappears as the yolk stalk obliterates and the amnion fuses with the umbilical cord.
The peritoneal cavity is continuous with a large chamber in the proximal part of the umbilical cord called the umbilical coelom, which surrounds the herniated portion of the midgut loop.
B. PLEURAL AND PERICARDIAL CAVITIES (THORACIC PORTION)
As each primitive lung sac expands, it encroaches on the pericardioperitoneal canal converting the canal into a primitive pleural cavity. On each side the cavity is separated from the more ventral pericardial cavity by the pleuropericartfial membrane. This membrane begins as a fold of tissue that is pulled from the lateral body wall to the mesocardium by the common cardinal veins when the heart migrates caudally. The membrane gradually narrows the pleuropericardial opening.
Each primitive pleural cavity communicates with the peritoneal cavity through the pleuroperitoneal opening. A fold called the pleuroperitoneal membrane forms between the mesonephros and the septum transversum and begins to close the pleuroperitoneal opening.
As the primitive lung sacs and body wall expand the primitive pleural cavities increase in size.
VI. MESENTERIES
A. DORSAL MESENTERY
The dorsal mesentery becomes thinner and suspends the gut from the dorsal body wall. It attaches to the gut from the caudal part of the esophagus to the caudal part of the hindgut near the rectum.
The dorsal mesogastrium attached to the greater curvature of the stomach moves to the left with the greater curvature. As a result, part of the peritoneal cavity called the lesser sac or omental bursa is isolated dorsal to the stomach. The rest of the peritoneal cavity is known as the greater sac. Part of the lesser sac extends toward the right lung bud as the pneumatoenteric recess.
The dorsal mesoduodenum is pulled to the right of the midline when the duodenum moves to the right. It receives the ventral pancreas.
The dorsal mesentery of the midgut loop twists in a counterclockwise direction along with the midgut but remains attached to the dorsal body wall.
The dorsal mesentery of the hindgut is short, broad and lies in the midline.
B. VENTRAL MESENTERY
A ventral mesentery attaches the terminal segment of the esophagus, lesser curvature of the stomach and adjacent part of the duodenum to the ventral body wall.
The liver grows at such a rapid rate that it expands into the ventral mesentery subdividing it into two portions; the portion between the gut and the liver is called the lesser omentum, the portion between the liver and the ventral body wall is called the falciform ligament. As the liver expands into the ventral mesentery, it is covered by a thin layer called the serosa. That part of the liver adjacent to the septum transversum has no such covering and is called the bare area.
Blood vessels course through the caudal edge of both subdivisions of the ventral mesentery; the portal vein, hepatic artery and bile duct course through the edge of the lesser omentum, the umbilical vein passes through the edge of the falciform ligament.
As a result of the stomach moving to the left and the duodenum to the right, the lesser sac opens to the right into the greater sac through a narrow opening called the epiploic foramen. This passageway lies dorsal to the portal vein.
VII. CARDIOVASCULAR SYSTEM
A. HEART
Figs. 6–4, 6–5GENERAL
The heart is located primarily in the cervical segments. The mesocardium is separated into cranial and caudal parts by a gap that will produce the transverse sinus in the pericardial cavity.
As the epimyocardial space disappears, three layers become apparent: a) the endocardium or thin lining of the heart chambers, b) the myocardium or thick muscular layer and c) the epicardium or thin surface layer.
SEPTUM FORMATION IN THE TRUNCUS ARTERIOSUS AND AORTIC SAC
A pair of opposing ridges called right (inferior) and left (superior) truncus swellings develops in the form of a spiral in the wall of the truncus arteriosus.
These two swellings grow toward one another, meet and fuse, thus forming the truncus septum that divides the lumen of the truncus arteriosus into two vessels, the ascending aorta and the pulmonary trunk. Because of the spiral arrangement of the swellings, the two vessels twist upon one another.
The area where the first four pairs of aortic arches arise from the aortic sac moves to the right. The point where the sixth pair of aortic arches arises from the sac moves to the left and becomes separated from the other arches by the thick aorticopulmonary septum. This septum is continuous with the spiral truncus septum thus causing the pulmonary trunk to course to the sixth aortic arches and the ascending aorta to course to the remaining arches.

FIG. 6–4
Heart of the 10-mm embryo. [printable version of FIG 6-4]
SEPTUM FORMATION IN THE CONUS CORDIS
Larger but slower developing swellings appear in the wall of the conus cordis. The right conus swelling forms along the right dorsal wall and becomes continuous with the left truncus swelling. The left conus swelling forms along the left ventral wall and becomes continuous with the right truncus swelling.
As the conus swellings grow toward one another, they fuse to form the conus septum that divides the conus into two portions: a) a ventrolateral portion that joins with the primitive right ventricle thereby forming the outflow tract of the definitive right ventricle and b) a dorsomedial portion that joins the primitive left ventricle thus forming the outflow tract of the definitive left ventricle.
SEPTUM FORMATION IN THE ATRIOVENTRICULAR (A-V) CANAL
As the bulboventricular flange recedes, the atrioventricular canal enlarges and moves to the right. Atrial blood then passes directly into both ventricles.
Two mesenchymal proliferations called endocardial cushions develop in the wall of the atrioventricular canal. The ventral (superior) cushion projects from the ventrocranial wall of the canal; the dorsal (inferior) cushion projects from the dorso-caudal wall. As the two cushions meet and fuse, they divide the previously single canal into two canals, one on the right and one on the left. Two other smaller proliferations called lateral cushions project from the right and left walls of the canal.
The right conus swelling becomes continuous with both the right lateral and dorsal cushions on the right side of the atrioventricular canal.
SEPTUM FORMATION IN THE VENTRICLES
The walls of the two ventricles expand in a caudal direction as the myocardium grows at a rapid rate. Muscular bands called trabeculae carneae cordis are produced in the inner part of the myocardium.
As the ventricles expand, their walls become adjacent caudal to the secondary interventricular foramen where they form the muscular portion of the interventricular septum.
Proliferation of the dorsal atrioventricular cushion reduces the size of the secondary intraventricular foramen.
SEPTUM FORMATION IN THE ATRIUM AND INCORPORATION
OF THE RIGHT SINUS HORN AND COMMON CARDINAL VEIN
As both portions of the atrial chamber continue to expand, the septum primum lengthens and approaches the endocardial cushions. This gradually narrows the ostium primum and better defines the boundary between the right and left atria.
A common pulmonary vein from the region of the branching lung buds courses through the caudal part of the mesocardium to join the left atrial wall.
The right horn of the sinus venosus and the right common cardinal vein continue to enlarge and become incorporated into the right atrium. This pulls the narrow transverse portion of the sinus venosus along with the left sinus horn and common cardinal vein to a position dorsal to the heart in the atrioventricular sulcus.
With the formation of the left venous valve, the sinoatrial orifice then has a valve on each side. Cranial to the orifice the venous valves join together to produce a ridge called the septum spurium. The bay area between the septum spurium and septum primum is called the septovalvular space.
The joining of the right sinus horn to the right atrium and the shifting of the venous return to the right side cause the right atrium to dilate and bulge to the right. This produces an infolding in the roof of the septovalvular space giving rise to the septum secundum.
B. ARTERIES
Fig. 6–5AAORTIC ARCHES
As the sixth pair of aortic arches develops, the first and second pairs disappear. The distal segment of the right sixth arch and the segment of the right dorsal aorta between the seventh intersegmental artery and the point of fusion of the two dorsal aortas begin to narrow and eventually disappear. According to some accounts a fifth pair of aortic arches develops but is very transient.
The origins of the arches from the aortic sac realign themselves in such a way that the third and fourth pairs arise from horns of the ventral part of the sac and become continuations of the ascending aorta. The sixth pair arises from the dorsal part of the sac and becomes a continuation of the pulmonary trunk.
The proximal part of the internal carotid artery is a remnant of the third arch. Its distal part is derived from the cranial segment of the dorsal aorta. The external carotid artery arises as a new branch from the ventral portion of the third arch. The segment of the dorsal aorta between the third and fourth arches begins to narrow and is called the carotid duct. It eventually disappears. The sixth pair of arches gives rise to a pulmonary artery on each side near the pulmonary trunk. Each artery courses to its respective lung bud.
Near the brain the internal carotid artery gives branches to the eye and diencephalon and then terminates as anterior and middle cerebral arteries to the telencephalon. The anterior cerebral artery will join its counterpart on the other side to form the anterior communicating artery across the midline.
The dorsal intersegmental arteries in the cervical segments anastomose into a longitudinal channel called the vertebral artery. Ventral to the myelencephalon the vertebral artery on each side joins its counterpart in the midline to form the single basilar artery. The basilar artery supplies the myel- and metencephalon before it bifurcates near the mesencephalon into right and left posterior cerebral arteries that supply the di- and mesencephalon and later on the telencephalon. An anastomosis called the posterior communicating artery develops on each side connecting the posterior cerebral and internal carotid arteries. A circular vascular channel called the Circle of Willis is thereby completed at the base of the brain.
Just cranial to the fusion point each dorsal aorta gives rise to an axial artery to the upper limb bud. The axial artery begins as a bud from the seventh cervical intersegmental artery.
COMMON DORSAL AORTA (AORTA)
The point of fusion of the paired dorsal aortas into a single aorta moves from the midcervical to the lower cervical segments.
The celiac, superior mesenteric and inferior mesenteric arteries become distinct, single vessels arising from the ventral aspect of the dorsal aorta. With the disappearance of the yolk stalk the distal part of the original vitelline artery eventually disappears. The proximal segment becomes the superior mesenteric artery that supplies the derivatives of the midgut.
Numerous lateral segmental branches of the dorsal aorta course to the mesonephros as mesonephric arteries.
The dorsal aorta bifurcates in the L-4 segment into right and left common iliac arteries each of which, after giving rise to the axial artery of the lower limb bud, continues into the umbilical cord as an umbilical artery. The axial artery develops as a bud of the fifth lumbar intersegmental artery.

FIG. 6–5
Cardiovascular system of the 10-mm embryo. [printable version of FIG 6-5]
- Arteries.
- Veins.
B. VEINS
Fig. 6–5BVITELLINE VEINS
The anastomotic network formed between the paired vitelline veins around the duodenum becomes a single channel that courses first to the left and then dorsal to the duodenum. This channel lies between the dorsal and ventral pancreas and represents the portal vein.
With the disappearance of the yolk stalk the distal tributaries of the vitelline veins eventually disappear. The proximal tributaries unite to form the superior mesenteric vein, which drains the midgut derivatives. The superior mesenteric vein courses through the dorsal mesentery to the pancreas area where it receives the splenic vein. It then continues to the liver as the portal vein.
As the left hepatocardiac vein disappears, blood from the left side of the liver is diverted to the right side. This causes dilation of the right hepatocardiac vein, which empties into the right horn of the sinus venosus.
UMBILICAL VEINS
An anastomosis forms between the proximal segment of each umbilical vein and the hepatic sinusoids. This segment of each umbilical vein is soon surrounded by the rapidly expanding liver cords and becomes incorporated into the liver, thus causing all of the umbilical blood to course through the liver sinusoids.
The entire right umbilical vein disappears forcing all of the chorionic blood to flow to the liver through a single channel, the former left umbilical vein.
As blood flow through the liver increases, a large channel called the ductus venosus develops as a direct communication between the umbilical vein and the right hepatocardiac vein thus allowing umbilical blood to bypass the hepatic sinusoids in its course to the heart. Blood in the portal vein also empties into the ductus venosus.
Some blood continues to enter the hepatic sinusoids by way of afferent veins, which arise from the ductus venosus. Sinusoidal blood drains by way of efferent veins into the right hepatocardiac vein.
CARDINAL VEINS
Common Cardinal Veins
The left common cardinal vein narrows and moves to the right in the dorsal atrioventricular sulcus as the left sinus horn is pulled to the right.
The right sinus horn becomes incorporated into the wall of the right atrium. The right common cardinal vein will become the terminal segment of the superior vena cava.
Precardinal Vein
The precardinal vein is a continuation of the large primary head vein that receives the primitive maxillary vein and the anterior, middle and posterior cerebral plexuses.
It narrows as it passes through the cervical region where it receives the marginal vein from the upper limb bud.
Postcardinal Vein
The cranial segment of the postcardinal vein begins to narrow as the subcardinal vein increases in size.
The caudal segment of the postcardinal vein receives the marginal vein from the lower limb bud and anastomoses in the midline with its counterpart on the other side.
Subcardinal Vein
The subcardinal vein on each side enlarges and courses longitudinally along the medial aspect of the mesonephros and gonadal ridge.
The right subcardinal vein establishes a connection with the right hepatocardiac vein.
The right subcardinal vein anastomoses across the midline with its counterpart on the other side.
COMMON PULMONARY VEIN
As the septum primum divides the common atrium into two chambers, veins from the region of the branching lung buds join together and enter the left atrium as a common pulmonary vein.
LYMPHATICS
Lymphatic channels begin as spaces or clefts in the mesenchymal tissue. A particularly large space develops in the cervical region called the jugular lymph sac which joins the precardinal vein.
VIII. SKELETAL SYSTEM
A. SKULL
Morphogenesis of the skull is divided into two parts: the neurocranium, which encloses the brain, and the viscerocranium, which supports the face, pharynx and larynx. The skull first appears as mesenchymal condensations which later either form membranes that ossify or first become cartilage that secondarily ossifies.
NEUROCRANIUM
The cartilaginous portion of the neurocranium (chondrocranium) forms the base of the skull. It is represented first by the following mesenchymal collections:
-
On each side of the cranial segment of the notochord constituting the parachordal condensation that blends caudally with the occipital sclerotomes.
Around the otocyst forming the otic capsule condensation.
Around the nasal pit forming the nasal capsule condensation.
VISCEROCRANIUM
The viscerocranium develops primarily from the branchial arch cartilages. These cartilages are represented first by mesenchymal condensations in the following regions:
-
First arch cartilage (Meckel’s)—medial to the mandibular nerve.
Second arch cartilage (Reichert’s)—medial to the facial nerve.
Laryngeal condensation—surrounding the primitive larynx and representing the caudal branchial arch cartilages.
B. POSTCRANIAL PART OF AXIAL SKELETON
The sclerotomes in each segment join around the notochord ventral to the neural tube.
Each sclerotome is composed of a caudal condensed part and a cranial less condensed part. The condensed part takes a position in the middle of each body segment with a spinal nerve on each side. The less condensed part is in the intersegmental position at the level of the dorsal intersegmental artery.
The sclerotomes from the occipital somites lose their segmentation and blend together around the notochord ventral to the myelencephalon. They fuse with the caudal part of the parachordal condensation and contribute to the formation of the occipital bone.
Caudal to the occipital region, cells from the cranial part of each sclerotome extend dorsally between the spinal ganglion and the myotome forming the neural arch.
Dense cellular condensations that develop from the twelve thoracic sclerotomes extend ventrolaterally as rib primordia.
IX. MUSCULAR SYSTEM
A. HEAD MESODERM
DORSAL TO THE OPTIC CUP
Premuscle masses for the extraocular muscles appear dorsal to the optic cup. They receive the terminal fibers of cranial nerves III, IV or VI. Although there is phylogenetic evidence that three preotic somites give origin to the extraocular muscles, they never form in human embryos. These muscles apparently differentiate in situ from mesenchyme.
BRANCHIAL ARCHES
Mesenchymal cells within each branchial arch rapidly increase in quantity. Some begin to collect around the terminal portion of the branchial arch nerves and represent the premuscle masses from which the branchial musculature develops.
The sternocleidomastoid-trapezius premuscle mass appears just caudal to the cervical sinus and already receives the spinal accessory nerve. Its origin is controversial, but it is derived probably from the mesenchyme in the caudal arches and from the cervical myotomes.
FLOOR OF THE PHARYNX
Premuscle masses of the tongue are evident in the mesenchyme ventral to the pharynx and are closely associated with the termination of the hypoglossal nerve. Although there is phylogenetic evidence that the occipital myotomes give origin to the tongue muscles, there is no continuity in human embryos between them and the tongue premuscle masses. The tongue muscles probably differentiate in situ from the mesenchyme in the floor of the pharynx.
B. MYOTOME
The occipital myotomes fuse into a single mass of myoblasts lateral to the proximal part of the hypoglossal nerve. The fate of this mass is unclear.
Caudal to the occipital region each myotome can be separated into a small, dorsal epiaxial division and a large ventrolateral hypaxial division. This is especially evident in the cervical region. The epiaxial divisions will give rise to the extensor muscles of the vertebral column in the back, which are innervated by the dorsal primary rami of spinal nerves. The hypaxial divisions will give rise to the lateral and ventral flexors of the vertebral column, which are innervated by the ventral primary rami of spinal nerves.
C. LATERAL (PLATE) MESODERM
The somatic layer between the mesothelial lining of the coelomic cavity and the ectoderm becomes thicker but is without distinct premuscle masses.
The splanchnic layer thickens and condenses around the gut and tracheal endoderm. It will form the muscularis of the alimentary tract and the cartilages in the wall of the tracheobronchial tree.
D. LIMB MESODERM
Limb mesoderm becomes very dense especially near the base and in the central part of the limb bud. Condensations begin to appear near the terminal ends of the spinal nerves.
The musculocutaneous, median, ulnar and radial branches of the brachial plexus can be identified as they pass into the upper limb bud.
Mesoderm in the lower limb bud receives the terminal nerve fibers of the lumbar and sacral plexuses. Identification of specific nerves is more difficult.
X. INTEGUMENTARY SYSTEM
A. ECTODERM
The surface of the embryo retains the single layer of ectoderm.
B. DERMATOME
The epitheloid character of the dermatome disappears as a result of cellular proliferation. Small islands of epitheloid cells are dermatome remnants sometimes present in the more caudal segments.
As the dermatome breaks up, many mesenchymal cells are produced, which spread beneath the expanding ectoderm.
C. MAMMARY GLAND
A thickened band of ectoderm called the mammary ridge becomes evident along the lateral body wall. It extends in a longitudinal manner from the upper to the lower limb bud (Fig. 6–1A).
XI. SPECIAL SENSE ORGANS
A. EYE
Fig. 6–1OPTIC VESICLE
The optic vesicle folds in on itself to become a double-layered optic cup surrounded by loose mesenchyme. The intraretinal space separates the inner and outer layers.
The outer layer is thin and becomes the pigmented layer of the retina.
The thick inner layer surrounds the optic cup cavity and becomes the visual layer of the retina. It differentiates into three zones that are similar to the neural tube wall called the ependymal, mantle and marginal layers. The deepest zone is the ependymal layer, which is separated from the pigmented layer of the retina by the intraretinal space.
The ventral surface of the optic cup and adjacent stalk folds inward to form the choroid fissure through which loose mesenchyme passes into the cup cavity.
LENS PLACODE
The lens placode invaginates to form the hollow lens vesicle and lens cavity. The lens vesicle loses contact with the ectoderm and lies within the cavity of the optic cup. The overlying ectoderm will contribute to the formation of the cornea.
The epithelial cells in the deep wall of the lens vesicle (deep lens epithelium) lengthen and project into the lens cavity. They will give rise to the entire lens with the exception of the superficial (anterior) lens epithelium that develops from the superficial wall.
EXTRAOCULAR PREMUSCLE MASSES
Premuscle masses for the extraocular muscles become evident dorsal to the optic cup and receive the terminal fibers of cranial nerves III, IV or VI.
B. EAR
INTERNAL EAR
Figs. 6–2A, 6–5BThe medial wall of the otocyst evaginates dorsally to produce the endolymphatic diverticulum. The diverticulum terminates near the roof of the fourth ventricle.
The otocyst constricts into a large dorsal portion called the vestibular pouch and a narrow, tapered ventral portion called the cochlear pouch. The lateral groove that appears on the lateral surface of the vestibular pouch demarcates the posterior semicircular canal.
The acoustic part of the facioacoustic neural crest becomes the vestibulocochlear nerve (VIII). Ganglion cells begin to differentiate in the area of contact between the nerve and the otocyst. The vestibular ganglion forms adjacent to the vestibular pouch and the cochlear (spiral) ganglion develops adjacent to the cochlear pouch.
Mesenchymal cells begin to accumulate around the otocyst forming the otic capsule condensation.
MIDDLE EAR
Fig. 6–2BThe first pharyngeal pouch extends dorsally as the tubotympanic recess and makes contact with the first branchial groove. The tissue in the area of contact represents the primitive tympanic membrane.
The first (Meckel’s) and second (Reichert’s) arch cartilages are present as precartilage condensations medial to the mandibular and facial nerves, respectively. The lateralmost extent of the tubotympanic recess lies between these nerves. The geniculate ganglion of the facial nerve differentiates just above the recess and the chorda tympani branch passes just below.
EXTERNAL EAR
Fig. 6–1ASmall elevations called auricular hillocks become evident on the surface of the first and second branchial arches adjacent to the first branchial groove. Three appear on each arch.
C. OLFACTORY APPARATUS
Fig. 6–1ATHE 10-MM EMBRYO
STAGE 16
AGE 37 DAYS
CROWN TO RUMP LENGTH 10.5 MM
Carnegie collection 6517
Reference
Streeter GL: Developmental horizons in human embryos. Description of age groups XV, XVI, XVII and XVIII. Contrib Embryol Carnegie Instn 32:133–203, 1948
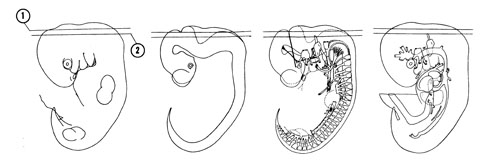
FIG. 6–6
SECTION 1
A section through the roof of the fourth ventricle and origin of the trochlear nerve.
Observe:
- The dorsal edge of the met- and myelencephalic parts of the rhombencephalon.
- The large fourth ventricle with its thin roof.
- The ventricle of the caudal part of the mesencephalon.
A section through the mesencephalic isthmus and tip of the endolymphatic diverticulum.
Observe:
- The alar plate of the myelencephalon with the posterior cerebral venous plexus on its surface.
- The alar and basal plates of the metencephalon separated by the sulcus limitans.
- The rhombic lip of the alar plate where the cerebellum will later develop.
- The minute trochlear nerve coursing ventrally on each side of the isthmus.

FIG. 6–6 [printable version of FIG 6-6]
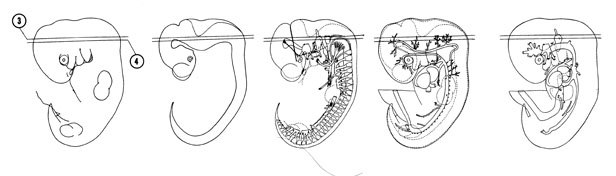
FIG. 6–7
SECTION 3
A section through the middle of the fourth ventricle and rootlets of the vagus and cranial accessory nerves.
Observe:
- The three parts of the undulating sulcus limitans that separates the alar and basal plates.
- The three distinct layers of the neural tube.
- The dorsal tip of the vestibular pouch that will form part of the internal ear.
A section through the region of the cephalic flexure, rootlets of the oculomotor nerve and origin of the glossopharyngeal nerve.
Observe:
- The cranial and spinal parts of the accessory nerve and the superior ganglion of the vagus nerve.
- The junction of the endolymphatic diverticulum with the medial wall of the vestibular pouch.
- The lateral groove in the lateral wall of the vestibular pouch which is surrounded by the otic capsule condensation.
- The three subdivisions of the mesencephalon: tectum, tegmentum and basis pedunculi area.

FIG. 6–7 [printable version of FIG 6-7]
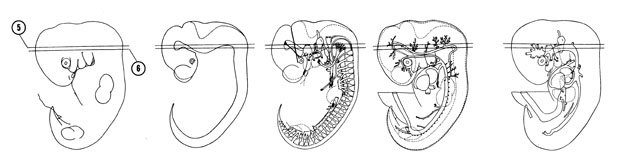
FIG. 6–8
SECTION 5
A section through the rootlets of the facial, vestibulocochlear and hypoglossal nerves.
Observe:
- The pyramidal tract area on the ventral side of the myelencephalon.
- The rhombomeres in the wall of the fourth ventricle.
- The union of the cranial accessory nerve with the vagus nerve.
- The superior ganglion of the glossopharyngeal nerve.
A section through the region of the cervical flexure, trigeminal nerve root and mamillary area of the diencephalon.
Observe:
- The cranial edge of the occipital myotomes.
- The anterior, middle and posterior cerebral plexuses, which drain into the primary head vein.
- The vertebral arteries about to join to form the basilar artery which is continuous with the posterior cerebral artery.
- The vestibular ganglion of the vestibulocochlear nerve adjacent to the vestibular pouch.
- The trigeminal ganglion adjacent to the pons area of the metencephalon.

FIG. 6–8 [printable version of FIG 6-8]
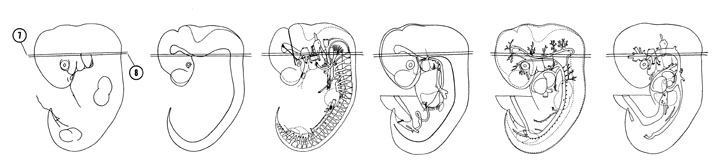
FIG. 6–9
SECTION 7
A section through the myelencephalon and spinal cord junction and the di- and mesencephalon junction of the neural tube.
Observe:
- The cochlear (spiral) ganglion of the vestibulocochlear nerve adjacent to the cochlear pouch.
- The geniculate ganglion of the facial nerve.
- The motor root of the trigeminal nerve medial to the trigeminal ganglion.
- The ventral edge of the fourth ventricle (median sulcus).
A section through the C-1 spinal ganglion, ventral edge of the pons area and neurohypophyseal bud.
Observe:
- The fused occipital sclerotomes that contribute to skull formation around the hypoglossal nerve.
- The inferior ganglion of the glossopharyngeal nerve.
- The close relation of the primary head vein to cranial nerves IX, X and XI and the trigeminal ganglion.
- The dorsal part of the first branchial groove in contact with the first pharyngeal pouch in the ear area.
- The sulcus medius between the dorsal thalamus and the ventral thalamus and the sulcus ventralis between the ventral thalamus and hypothalamus.

FIG. 6–9 [printable version of FIG 6-9]
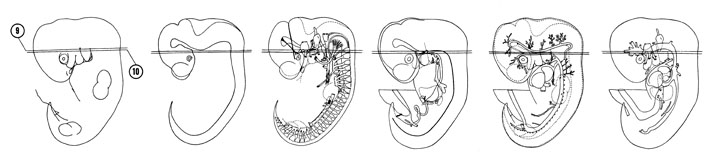
FIG. 6–10
SECTION 9
A section through the junction of the hypophyseal pouch with the neurohypophyseal bud.
Observe:
- The three divisions of a somite: sclerotome, myotome and dermatome.
- The basilar artery that travels in the midline on the ventral surface of the pons area in Section 8.
- The abducens nerve passing to the lateral rectus premuscle mass.
- The facial nerve in the middle of the second branchial arch.
- The three divisions of the trigeminal nerve; ophthalamic, maxillary and mandibular nerves.
A section through the cranial end of the notochord and edge of the primitive pharynx and internal carotid artery.
Observe:
- The four plate areas of the spinal cord.
- The anterior cerebral plexus and primitive maxillary vein that join with the primary head vein seen in Section 9.
- The caudal continuation of the primary head vein as the precardinal vein.
- The extraocular premuscle mass where cranial nerves III and IV terminate and the lateral rectus premuscle mass where cranial nerve VI terminates.
- The communication between the C-1 spinal nerve and the hypoglossal nerve.

FIG. 6–10 [printable version of FIG 6-10]
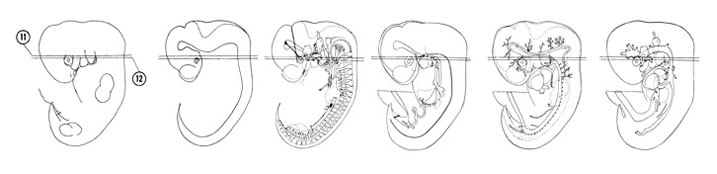
FIG. 6–11
SECTION 11
A section through the C-2 spinal ganglion and dorsal surface of the tongue.
Observe:
- The inferior ganglion of the vagus medial to the spinal accessory nerve.
- The maxillary and mandibular nerves in the first branchial arch.
- The facial and glossopharyngeal nerves on each side of the second pharyngeal pouch.
- The chorda tympani nerve arising from the facial nerve.
- The internal carotid artery flanking each side of the hypophyseal pouch.
A section through the premuscle mass of the tongue and the junction of the hypophyseal pouch with the stomodeum.
Observe:
- The spinal accessory nerve in the sternocleidomastoid-trapezius premuscle mass.
- The epiglottis projecting into the primitive pharynx.
- The termination of the glossopharyngeal nerve near the tongue surface.
- The opercular fold and cartilage of the second branchial arch.
- The optic cup, stalk and chiasmal area of the hypothalamus.

FIG. 6–11 [printable version of FIG 6-11]
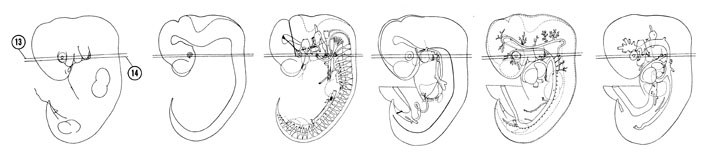
FIG. 6–12
SECTION 13
A section through the optic stalk lumen and lens vesicle.
Observe:
- The dense mesenchyme of the second branchial arch where the facial nerve has terminated.
- The dorsal portion of the third pharyngeal pouch where the caudal (inferior) parathyroid gland develops.
- The parts of the lens vesicle and layers of the optic cup.
- The three layers of the spinal cord.
- The thyroglossal duct in the midline between the primitive pharynx and tongue musculature.
A section through the C-3 spinal ganglion, branchial arches and thyroid gland.
Observe:
- The cervical sinus ventral to the sternocleidomastoid-trapezius premuscle mass.
- The ventral portion of the third pharyngeal pouch which gives rise to the greater thymic bud.
- The termination of the hypoglossal nerve in the premuscle masses of the tongue.
- The first branchial arch cartilage in the mandibular process.
- The superior laryngeal branch of the vagus nerve that innervates part of the larynx

FIG. 6–12 [printable version of FIG 6-12]
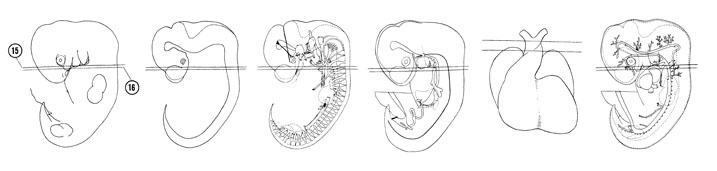
FIG. 6–13
SECTION 15
A section through the primitive larynx and fourth pharyngeal pouch.
Observe:
- The dorsal and ventral primary rami of the C-3 spinal ganglion.
- The dorsal portion of the fourth pharyngeal pouch that gives rise to the rostral (superior) parathyroid gland and the ventral portion that forms the lesser thymus.
- The arytenoid swelling on each side of the primitive larynx.
- The lamina terminalis and the di- and telencephalon junction.
- The maxillary process and lateral nasal elevation separated by the maxillonasal groove.
A section through the C-4 spinal ganglion and the cranial edge of the heart.
Observe:
- The separation of the pharyngeal endoderm into esophageal and tracheal portions.
- The infrahyoid premuscle mass on either side of the heart in the cervical region.
- The right and left atria separated by the ascending aorta and pulmonary trunk.
- The edge of the nasal pit with the nasal fin separating the medial nasal elevation from the maxillary process.
- The two subdivisions of the telencephalon: cerebral vesicle and telencephalon medium.

FIG. 6–13 [printable version of FIG 6-13]
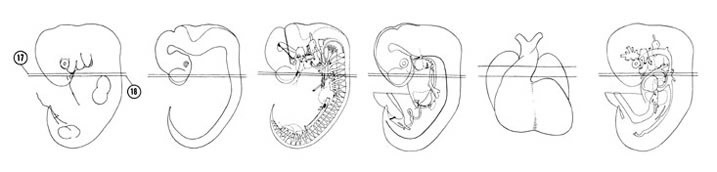
FIG. 6–14
SECTION 17
A section through the truncus arteriosus of the heart and cerebral vesicle of the telencephalon.
Observe:
- The dorsal and ventral primary rami of the C-4 spinal ganglion coursing to the epiaxial and hypaxial myotomic divisions, respectively.
- The right and left truncus swellings dividing the truncus arteriosus into the ascending aorta and pulmonary trunk.
- The junction of the truncus arteriosus and conus cordis.
- The medial and lateral nasal elevations on each side of the nasal pit.
- The two parts of the cerebral vesicle: striatal and suprastriatal.
A section through the C-5 spinal ganglion and the conus swellings of the heart.
Observe:
- The vagus nerve lateral to the esophagus and trachea.
- The pulmonary artery ventral to the trachea.
- The phrenic nerve lateral to the right atrium.
- The septum primum and septum secundum between the right and left atria.
- The olfactory fila between the olfactory epithelium and the primordial tuberculum olfactorium.

FIG. 6–14 [printable version of FIG 6-14]
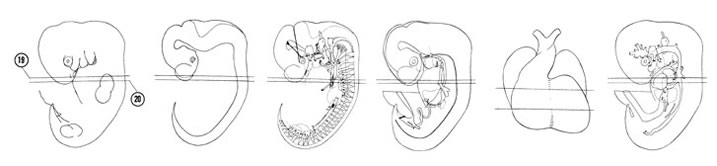
FIG. 6–15
SECTION 19
A section through the atrioventricular (AV) region of the heart.
Observe:
- The neural arch in the C-5–C-6 interganglion region.
- The junctions of the pre- and postcardinal veins and of the right common cardinal vein with the right sinus horn.
- The right and left venous valves between the right sinus horn and right atrium.
- The fusion of the endocardial cushions with the ostium primum above and the interventricular foramen below.
- The union of the medial and lateral nasal elevations.
A section through the C-6 spinal ganglion and tracheal bifurcation forming primary bronchi.
Observe:
- The brachial plexus surrounding the axial artery.
- The pleuropericardial fold separating the primitive pleural and pericardial cavities.
- The entrance of the common pulmonary vein into the left atrium.
- The three layers of the heart wall: epicardium, myocardium and endocardium.
- The trabeculae carneae cordis projecting into the ventricular chambers.
 region of the heart. A section through the C-6 spinal ganglion and tracheal bifurcation forming primary bronchi.")
FIG. 6–15 [printable version of FIG 6-15]
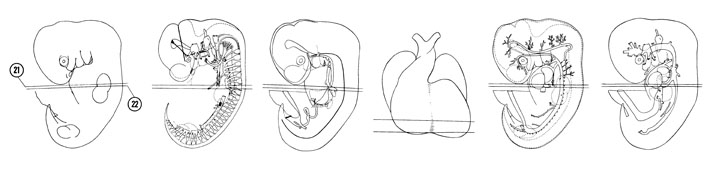
FIG. 6–16
SECTION 21
A section through the C-7 spinal ganglion and left sinus horn.
Observe:
- Three large brachial plexus nerves innervating the upper limb bud.
- The junction of the dorsal aortas into a common midline vessel.
- The junction of the left common cardinal vein with the left sinus horn.
- The edge of the liver separated from the pericardial cavity by the septum transversum.
- The muscular portion of the interventricular septum separating the ventricles.
A section through the pleuroperitoneal membrane and the cranial edge of the mesonephros.
Observe:
- The primitive lung sac bulging into the primitive pleural cavity.
- The separation of the primitive pleural and peritoneal cavities by the pleuroperitoneal membrane.
- The termination of the phrenic nerves in the septum transversum.
- The right hepatocardiac vein that joins the right sinus horn in Section 21.
- The caudal edge of the right ventricle.

FIG. 6–16 [printable version of FIG 6-16]
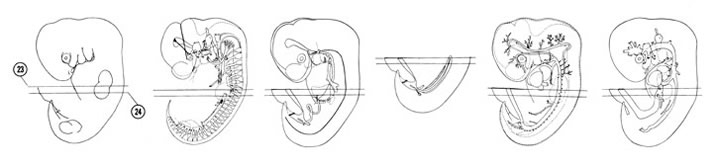
FIG. 6–17
SECTION 23
A section through the C-8 spinal ganglion and first rib primordium.
Observe:
- The primitive hand plate of the upper limb bud with the peripheral marginal vein.
- The communication of the primitive pleural and peritoneal cavities through the pleuroperitoneal opening ventral to the mesonephric duct.
- The cranial end of the mesonephric duct.
- The junctions of an efferent vein and the ductus venosus with the right hepatocardiac vein.
- The components of the umbilical cord.
A section through the T-1 spinal ganglion and stomach.
Observe:
- The dorsal and ventral mesogastrium attaching to the greater and lesser curvatures of the stomach, respectively.
- The lesser sac dorsal to the stomach.
- The junction of the portal vein with the ductus venosus.
- Nearby afferent veins arising from the ductus venosus carrying blood to the hepatic sinusoids.
- The edge of the gall bladder on the caudal surface of the liver.

FIG. 6–17 [printable version of FIG 6-17]
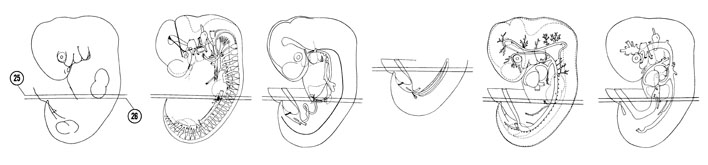
FIG. 6–18
SECTION 25
A section through the T-2 spinal ganglion, suprarenal gland and gastroduodenal junction.
Observe:
- A ramus communicans connecting the sympathetic trunk with a ventral primary ramus.
- The splenic primordium in the dorsal mesogastrium.
- The lesser sac communicating through the epiploic foramen with the greater sac.
- The portal vein, bile duct and hepatic artery ventral to the epiploic foramen.
- The junction of the ductus venosus and umbilical vein at the liver surface.
A section through the T-3 spinal ganglion and caudal edge of the liver.
Observe:
- A mesonephric glomerus within a capsule.
- The cranial end of the gonad.
- The close relation of the celiac plexus to the suprarenal medulla in Section 25.
- The superior mesenteric vein between the dorsal and ventral pancreas.
- The junction of the dorsal pancreas with the duodenum.

FIG. 6–18 [printable version of FIG 6-18]
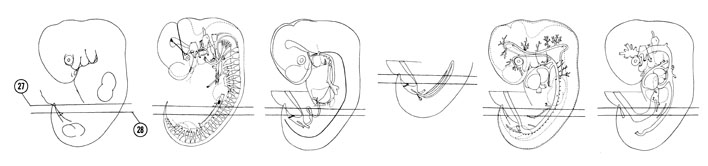
FIG. 6–19
SECTION 27
A section through the T-4 spinal ganglion, herniated midgut and tip of the tail bud.
Observe:
- The thickest part of the mammary ridge where the mammary gland develops.
- The extension of the peritoneal cavity into the umbilical cord.
- The duodenum and its mesentery to the right of the superior mesenteric artery and vein.
- The primitive filum terminale or tail segment of the neural tube.
- The narrow allantois between the umbilical arteries.
A section through the T-6 spinal ganglion and junction of the umbilical vessels with the ventral body wall.
Observe:
- The position of the intercostal nerve to the dorsal end of the rib primordium.
- The anastomosis between the post- and subcardinal veins medial to the mesonephros.
- The junction of a glomerular capsule and tubule in the mesonephros.
- The superior mesenteric vessels in the twisted dorsal mesentery of the midgut.
- The marginal vein and placode of the primitive foot plate.

FIG. 6–19 [printable version of FIG 6-19]
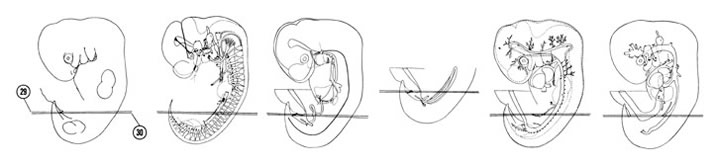
FIG. 6–20
SECTION 29
A section through the primitive cecum and genital tubercle.
Observe:
- The three layers of the spinal cord.
- The caudal edge of the cranial limb of the midgut loop to the right of the midline.
- The urethral plate on the ventral surface of the genital tubercle.
- The continuity of the urethral plate with the cloacal membrane in Section 30.
A section through the labioscrotal swelling and the urogenital groove and fold.
Observe:
- The coelomic bays of the peritoneal cavity medial and lateral to the mesonephric ridge.
- The junction of a tubule with the mesonephric duct.
- The apex of the urogenital sinus that is continuous with the allantois in Section 29.
- The platelike cloacal membrane in the depths of the urogenital groove.
- The junction of the tail bud with the body.

FIG. 6–20 [printable version of FIG 6-20]
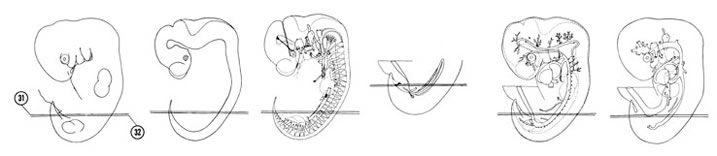
FIG. 6–21
SECTION 31
A section through the T-7 spinal ganglion.
Observe:
- The mesonephric duct coursing longitudinally on the lateral aspect of the mesonephros.
- The edge of the mesoderm that surrounds the midgut loop in Section 29.
- The mid- and hindgut junction.
- The caudal edge of the umbilical vein.
- The junction of the cloacal membrane with the cloaca.
A section through the urogenital sinus, rectum and urorectal septum.
Observe:
- The myotome lateral to the neural arch.
- The sympathetic trunk dorsolateral to the dorsal aorta.
- The thick gonadal ridge in the ventromedial portion of the mesonephric ridge.
- The hindgut suspended by a dorsal mesentery.
- The urorectal septum dividing the cloaca into the urogenital sinus and rectum.

FIG. 6–21 [printable version of FIG 6-21]
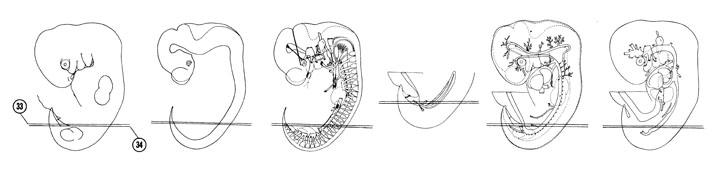
FIG. 6–22
SECTION 33
A section through the horns of the urogenital sinus near their junction with the mesonephric duct.
Observe:
- The splanchnic nerve arising from the thoracic part of the sympathetic trunk.
- The inferior mesenteric artery in the dorsal mesentery coursing to the hindgut.
- The rectum without a mesentery.
A section through the T-8 and coccygeal spinal ganglia.
Observe:
- The sympathetic neuroblasts surrounding the dorsal aorta.
- The two segments of the mesonephric duct.
- The junction of the hindgut with the rectum.
- The minute metanephric diverticulum near its origin from the urogenital sinus horn in Section 33.

FIG. 6–22 [printable version of FIG 6-22]
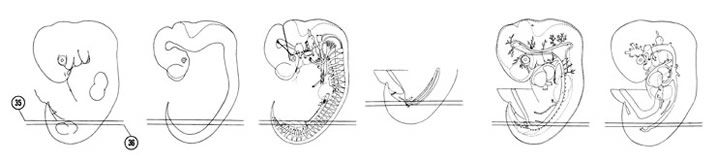
FIG. 6–23
SECTION 35
A section through the T-8 and S-5 spinal ganglia.
Observe:
- The junction of the two segments of the right mesonephric duct.
- The caudalmost extent of the mesonephros.
- The minute metanephric diverticulum.
A section through the T-9 and S-4 spinal ganglia.
Observe:
- A splanchnic nerve coursing from the sympathetic trunk to the dorsal aorta.
- The left metanephric diverticulum approaching the metanephrogenic mass.
- The blind dilated portion of the diverticulum called the primitive renal pelvis, which is surrounded by the metanephrogenic mass.
- The right common iliac artery is continuous with the right umbilical artery in Section 35.

FIG. 6–23 [printable version of FIG 6-23]
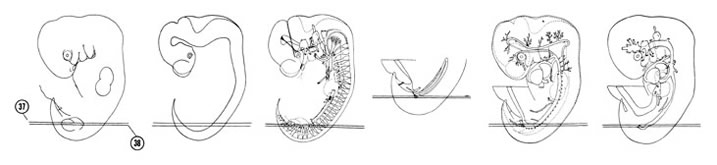
FIG. 6–24
SECTION 37
A section through the T-10 and S-3 spinal ganglia.
Observe:
- The spinal cord in oblique section.
- The bifurcation of the dorsal aorta in the L-4 segment into right and left common iliac arteries.
- The left metanephrogenic mass caudal to the bifurcation.
- The axial artery and marginal vein of the lower limb bud.
A section through the origin of the median sacral artery.
Observe:
- Sympathetic neuroblasts on the surface of the dorsal aorta.
- The sacral plexus in the proximal part of the lower limb bud.
- The dorsal edge of the metanephrogenic mass.

FIG. 6–24 [printable version of FIG 6-24]
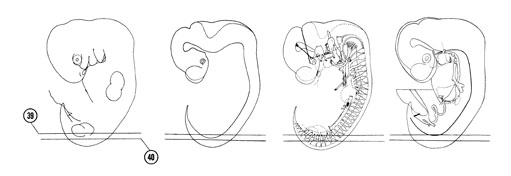
FIG. 6–25
SECTION 39
A section through the T-12 and S-2 spinal ganglia.
Observe:
- The sclerotomes with their loose rostral and dense caudal parts.
- The notochord in longitudinal section.
- The intersegmental position of the dorsal intersegmental arteries.
- The segmental position of the spinal nerves and myotomes.
- The dermatome cells between the ectoderm and the myotome.
A section through the L-1 to S-1 spinal ganglia.
Observe:
- The basal plate and neural canal in longitudinal section.
- The ventral roots of the lumbar spinal nerves emerging from the basal plate.
- The intersegmental position of the neural arches.
- The epitheloid remnants of the dermatome.

FIG. 6–25 [printable version of FIG 6-25]