raymond f. gasser
ATLAS OF HUMAN EMBRYOS
Copyright © 1975 RF Gasser, PhD. All rights reserved. Reproduced with permission.Chapter VIII (Pages 241-297)
- External Appearance
- Nervous System
- Alimentary and Respiratory Systems
- Urogenital System
- Coelomic Cavity
- Mesenteries
- Diaphragm
- Cardiovascular System
- Skeletal System
- Muscular System
- Integumentary System
- Special Sense Organs
the eighth week of life
late embryonic period
I. EXTERNAL APPEARANCE
Fig. 8–1AThe specimens measure approximately 21 to 30 mm in length.
The head region composes almost half of the embryo, its large size being due to the growth of the cerebral hemispheres. The face occupies only the lower one-half of the head. As the head continues to raise from the ventral chest wall, the cervical fold disappears and the neck region becomes well defined.
The eyes move into the frontal plane closer to the midline and nearly attain their definitive position. The eyelid folds lengthen and begin to cover the exposed surface of the eye. A short period later (35 mm) the folds will meet and fuse. They will remain fused until around the seventh month.
The auricle begins to assume its definitive shape around the external acoustic meatus.
The face exhibits smooth contours as the elevations and processes disappear. The nose becomes prominent around the external nostrils, which are adjacent to the midline and occluded by epithelial plugs. A distinct chin becomes evident in the midline at the ventral end of the mandibular area.
The upper limb bud can be called the upper limb or extremity as the shoulder, elbow and wrist areas become apparent. Likewise, the lower limb bud becomes the lower limb or extremity with the appearance of the knee and ankle areas. The limbs continue to rotate resulting in the palms and soles pointing caudally. This occurs to a greater degree in the upper limb than in the lower.
The large heart and liver bulge the ventral body wall but to a lesser degree than previously. The relative size of the umbilical cord is reduced. The infraumbilical area between the pubis and the umbilical cord begins to appear.
The surface features of the external genitalia (perineal region) are described in Section IV, F.
All evidence of a tail bud disappears.
II. NERVOUS SYSTEM
A. CENTRAL NERVOUS SYSTEM (CNS)
Fig. 8–1BBy the end of the second month some reflex activity is demonstrated. Lightly stroking the lips causes contralateral flexion of the neck and upper trunk. Some of the sensory and motor neurons therefore have functional connections both centrally and peripherally.
GENERAL
The brain continues to enlarge at a faster rate than the spinal cord. This is especially evident in the cerebral hemispheres (vesicles). As the head raises, the rhombencephalon assumes a more vertical position and the cervical flexure becomes less acute. The cephalic and pontine flexures become more acute. The spinal cord begins to lag in its ability to keep pace with the increasing length of the vertebral column. As a result, the caudal end of the spinal cord begins to move cranially in relation to the vertebral column. It remains attached to the coccygeal region by the primitive filum terminale.
The ectomeninx differentiates into an outer layer that has osteogenic or chondrogenic properties and an inner layer called the dura mater. Venous sinuses or plexuses separate the two layers in the cranium. In the vertebral column the two layers are separated by a venous plexus around which a large epidural space will subsequently develop. The endomeninx differentiates into the pia mater around the blood vessels on the surface of the CNS and the arachnoid trabeculae that attach to the deep surface of the dura mater. The arachnoid appears in late fetal life as a delamination from the deep aspect of the dura mater. When this occurs, a very narrow subdural space separates the dura mater and arachnoid and a broad subarachnoid space containing cerebrospinal fluid separates the arachnoid from the pia mater.
BRAIN
Telencephalon
-
Cerebral Vesicle (Cerebral Hemisphere)
Striatal Part
The cerebral vesicle expands at a rapid rate in a semicircular manner, growing dorsally first, then caudally. The striatal ridge in the floor of the lateral ventricle at the interventricular foramen also grows in a similar manner. The rostral part of the ridge divides into medial and lateral portions. The large basal ganglia subsequently differentiate from the striatal part of the cerebral vesicle.
The primordial piriform cortex at the surface is close to the primordial olfactory bulb where the olfactory nerves enter.
Suprastriatal Part
The lateral ventricle follows the dorsal, then caudal expansion of the cerebral vesicle and thereby produces an inferior horn. Likewise, the choroid fissure and plexus in the medial wall at the interventricular foramen are pulled in a similar manner.
The primordial hippocampus dorsal to the choroid fissure follows the same growth pattern lying first in the medial wall of the inferior horn, then in the floor. The amygdala area lies in the roof of the inferior horn between the rostral end of the horn and the basal ganglia. The wall of the telencephalon, in the area of the primordial olfactory bulb, evaginates to form the olfactory bulb. The ventricular cavity extends initially into the bulb but later disappears.
A thick layer of neuroblasts develops in the primordial neopallial cortex forming the superficial cortical layer of the neopallial cortex. Frontal, parietal, occipital and temporal lobe areas become apparent. Growth of the region between the frontal and temporal areas becomes depressed forming the insular area that lies lateral to the striatal part of the cerebral vesicle.
-
Telencephalon Medium
The preoptic recess is a distinct, isolated portion of the third ventricle and is bound on each side by the preoptic area. The preoptic area is separated from the hypothalamus by the optic groove and is delimited dorsally by the anterior commissure area where olfactory fibers cross the midline.
Diencephalon
-
Epithalamus
The epithalamus develops from the roof plate and the adjacent, dorsal part of the lateral wall. Its growth does not keep pace with the dorsal thalamus and it is reduced to a small region above the third ventricle.
The pineal bud becomes a distinct evagination of the roof plate and subsequently becomes the pineal gland (epiphysis).
The habenular area where the habenular nuclei develop is rostral to the pineal gland. The habenular commissure area appears as a clear area in the marginal layer in the midline. The posterior commissure area is similar in appearance caudal to the pineal gland.
-
Dorsal Thalamus
The dorsal thalamus enlarges greatly and makes up most of the dorsal part of the diencephalon. It lies dorsal to the hypothalamic sulcus, the formation of which is described below.
The internal capsule area is lateral. Caudally it is continuous with the mesencephalon.

FIG. 8 –1 [printable version of FIG 8-1]
- External features of the 30-mm embryo.
- Central nervous system of the 30-mm embryo. Arrows indicate the position of the cephalic, pontine and cervical flexures.
-
Ventral Thalamus
As the dorsal thalamus expands, the ventral thalamus moves from a position near the midline to one lateral to the hypothalamus.
As this occurs, the sulci medius and ventralis blend to form the hypothalamic sulcus, which continues rostrally into the preoptic recess as the optic groove.
-
Hypothalamus
The hypothalamus lies in the most ventral part of the diencephalic wall below the hypothalamic sulcus. Its rostral boundary is the optic chiasma; its caudal boundary is the mammillary area where the small, spherical mammillary body will develop on each side of the midline. The hypothalamus is continuous rostrally with the preoptic area of the telencephalon. Caudally it is adjacent to the tegmentum of the mesencephalon. The neuroblasts in the wall will give rise to the hypothalamic nuclei.
The neurohypophyseal bud gives rise to the neurohypophysis that is located in the midline just caudal to the optic chiasma. The tuberal part of the adenohypophysis covers it laterally.
Mesencephalon
-
Tectum
A longitudinal elevation called the collicular ridge develops in the roof plate. A transverse groove subsequently divides the ridge into superior and inferior colliculi that are relay centers of the visual and auditory senses, respectively.
The thick mantle layer gives rise to neuroblasts that migrate into the marginal layer in the form of sheets. This gives the tectum its stratified appearance.
-
Tegmentum
The neuroblasts in the tegmentum will differentiate into the motor nuclei of cranial nerves III and IV, the red nucleus and the substantia nigra. Those that remain dispersed contribute to the reticular formation.
The alar plate possibly contributes some neuroblasts to the tegmentum.
-
Basis Pedunculi Area
The basis pedunculi area located ventral to the tegmentum will be composed of fibers that originate primarily from the neopallial cortex.
It is continuous rostrally with the internal capsule area and caudally with the pons area.
Metencephalon
-
Pons Area
Basal Portion
Many of the fibers from the neopallial cortex will pass through the marginal layer making up the pyramidal tract area.
Some of these fibers will relay in the pons area and course to the cerebellum.
Tegmentum
The alar and basal plates are separated by a shallow sulcus limitans.
Neuroblasts thought to be from the alar plate of the myelencephalon extend rostrally into the pons area where they form the primordial superior olivary nucleus and, at a later time, the pontine nuclei.
-
Neuroblasts in the mantle layer of both the metencephalon and myelencephalon begin to segregate themselves into groups that become the nuclei of several cranial nerves. The motor nuclei are derived from the basal plate and the sensory nuclei are derived from the alar plate.
Motor nuclei—Three different columns of motor nuclei develop in the basal plate. Somatic motor nuclei differentiate medially near the midline; branchiomotor (special visceral motor) nuclei develop in the intermediate region, and general visceral motor nuclei form in the lateral part of the basal plate. The primordial abducens (somatic motor) and facial (branchiomotor) nuclei are among the first motor nuclei in the metencephalon to become apparent.
Sensory nuclei—Three different columns of sensory nuclei develop in the alar plate, their relative positions being a mirror image of the motor nuclei. General visceral sensory nuclei differentiate medially next to the sulcus limitans, special visceral sensory nuclei develop in the intermediate region and general somatic sensory nuclei form laterally next to the developing cerebellum. A fourth column called special somatic sensory nuclei is described in the adult. These nuclei lie between the special visceral and general somatic sensory nuclei and are relay centers for the vestibulocochlear nerve. The primordial nucleus of the spinal tract of V (general somatic sensory) and the rostral part of the primordial vestibular nucleus (special somatic sensory) are among the first sensory nuclei in the metencephalon to differentiate. The clear region lateral to the primordial nucleus of the spinal tract of V represents the spinal tract of V area.
-
Cerebellum
The intraventricular portion of the rhombic lip initially enlarges at a more rapid rate than the extraventricular portion and bulges into the fourth ventricle. The extraventricular will subsequently become larger than the intraventricular portion. Together they will form the cerebellar hemispheres.
In the rostral part of the metencephalon the rhombic lips will join together in the midline where the cerebellar vermis develops.
Medulla Oblongata (Myelencephalon)
All of the motor and sensory nuclear columns of the metencephalon are continuous into the medulla oblongata and retain their same relative positions. The roof plate narrows in the caudal part of the medulla oblongata bringing the alar plate and the related sensory nuclei into a dorsolateral position.
Basal Plate
The primordial hypoglossal nucleus (general somatic motor) becomes evident in the floor of the fourth ventricle next to the midline.Alar Plate
A large part of the primordial vestibular nucleus develops in the special somatic sensory nuclear column in the rostral portion of the medulla oblongata. Several primordial nuclei become apparent in the general somatic sensory nuclear column in the caudal portion of the medulla oblongata. From medial to lateral they are the primordial nucleus gracilis, nucleus cuneatus and nucleus of the spinal tract of V. A large number of neuroblasts thought to be derived from the alar plate collect in the ventral portion of the medulla oblongata to form the primordial inferior olivary and arcuate nuclei, both of which will send fibers to the cerebellum.Tract Areas
Several major tracts become evident as clear, less cellular areas. The pyramidal tract area is well defined in the ventral portion of the medulla oblongata. It will be composed of fibers from the neopallial cortex that descend to the motor nuclei in the spinal cord. Next to the midline the medial lemniscus area becomes evident and will contain primarily ascending fibers from the sensory nuclei in the spinal cord on their way to the thalamus. Many of the fibers cross to the opposite side in the sensory decussation area. The spinal tract of V area will contain descending sensory fibers from the skin of the face and auricle. It begins in the pons area just lateral to its primordial nucleus and will extend through the medulla oblongata. It will be continuous with a similar region in the dorsolateral part of the spinal cord (Lissauer’s tract).
Fourth Ventricle
The diamond shaped fourth ventricle narrows in the caudal portion of the medulla oblongata and becomes continuous with the neural (central) canal of the spinal cord.
The roof fuses with the vascular layer of the endomeninx (pia mater) thereby forming the tela choroidea. Folds of the tela project into the ventricle as the choroid plexus where cerebrospinal fluid will be produced. Cerebrospinal fluid will circulate throughout the brain ventricles. A midline outpocketing of the roof called the ependymal diverticulum communicates with the fourth ventricle through the median aperture. The diverticulum subsequently disappears after which the cerebrospinal fluid can pass from the ventricle into the subarachnoid space. The median aperture is then called the foramen of Magendie. A similar aperture called the foramen of Luschka will also appear in the roof on each side of the midline.
When the cerebellum enlarges it pushes the roof caudally and ventrally. Eventually the roof lies below the caudal part of the cerebellum.
SPINAL CORD
As the layers of the spinal cord expand, the neural canal is reduced to a tiny channel lined with ependyma, called the central canal. Its dorsal walls join in the midline to form the dorsal median septum. The central canal dilates caudally at the tip of the spinal cord into the terminal ventricle.
The neuroblasts are located primarily in the mantle layer, which gives rise to the grey matter. The marginal layer, containing predominantly nerve fibers, forms the white matter at the periphery of the grey matter. The grey matter arranges itself in the shape of a butterfly on cross section with dorsal and ventral horns or columns that arise from the alar and basal plates, respectively. A lateral horn develops in the thoracic and upper lumbar segments and is the seat of visceral motor nuclei. Sensory nuclei reside in the dorsal horn, somatic motor nuclei are located in the ventral horn. The dorsal and ventral roots of the spinal nerves divide the white matter into dorsal, lateral and ventral funiculi.
The ventral median sulcus on the brain is continuous onto the spinal cord with a deeper groove called the ventral median fissure. The ventral (anterior) spinal artery travels in the fissure. A shallower dorsal median sulcus appears in the midline on the dorsal surface.
B. PERIPHERAL NERVOUS SYSTEM
Because the vertebral column lengthens at a faster pace than the spinal cord, the spinal cord recedes cranially in the vertebral canal. In order to accommodate this differential growth pattern the spinal nerve roots attached to the caudal part of the cord must increase in length. As the tip of the spinal cord recedes progressively higher in the vertebral canal, the primitive filum terminale as well as progressively more spinal nerve roots increase in length. At birth the tip of the spinal cord is located at approximately the L-4 vertebra; in the adult it is located at approximately the L-2 vertebra. The many spinal nerve roots in the vertebral canal between the end of the spinal cord and the coccyx are collectively called the cauda equina (horse’s tail).
As the vertebral column develops, all but the most caudal spinal ganglia lie in the intervertebral foramina. Since the sacral foramina develop peripheral to the sacral spinal ganglia, these ganglia are located in the caudal part of the vertebral canal. In the adult the C-1 spinal ganglion is usually not apparent. The C-1 spinal nerve is predominantly motor.
All of the named peripheral nerves can be identified by the end of the eighth week. As viscera and muscle masses shift in their position, their nerves accommodate the shifts by increasing in length. The heart and diaphragm receive most of their innervation when they are in the cervical region. Since their nerves (superior, middle and inferior cervical cardiac nerves of the heart and phrenic nerve of the diaphragm) follow them as they migrate caudally, they begin in the neck and descend through the root of the neck and thorax. Likewise, the larynx receives its motor innervation from the vagus when the aortic arches are farther cranially. When the arches migrate caudally with the heart, the recurrent laryngeal nerve follows causing it to recur in its course to the larynx. In the adult it passes around the aortic arch and ligamentum arteriosum on the left, which were derived from the fourth and sixth arches, respectively. Since on the right side the distal part of the sixth arch disappears, the nerve loops around the proximal part of the subclavian artery that is derived from the fourth arch.
C. SUPRARENAL GLAND
The suprarenal gland is initially composed primarily of cortex with only a scant amount of medulla medially. The very large cortical mass lies on the dorsomedial surface of the definitive kidney. During late fetal life the medulla will be overgrown by the cortex causing it to occupy a more central position within the cortex. The cortex formed during the embryonic period becomes the fetal cortex, which disappears after birth. It is replaced later by the definitive cortex that is derived from smaller cells at the periphery.
III. ALIMENTARY AND RESPIRATORY SYSTEMS
Fig. 8–2AA. ORONASOFACIAL REGION
The lateral palatine processes move to a position above the tongue where they fuse in the midline with each other and with the lower edge of the nasal septum. The processes thereby become the secondary palate that completes the separation between the primitive nasal and oral cavities. These cavities are then called nasal and oral cavities. The caudalmost part of the secondary palate that gives rise to the soft palate and uvula is usually the last to form.
The primitive palate differentiates into the median part of the upper lip and jaw and the triangular shaped median palatine process. The lateral aspect of the median palatine process fuses on each side with the rostralmost segment of the lateral palatine process. The median palatine process becomes the premaxillary palate caudal to which is the incisive foramen. Here the median and two lateral palatine processes come together in a Y shaped manner.
The external nostril through which the nasal cavity communicates to the exterior is temporarily occluded by an epithelial plug. The nasal cavity is continuous with the nasopharynx through the internal nostril (choana). The medial wall of the nasal cavity contains the vomeronasal organ, which eventually disappears. The longitudinal elevations in the lateral wall represent the superior, middle and inferior concha (turbinates). The nasolacrimal duct terminates below the inferior concha.
The oral cavity communicates with the exterior through the rima oris. The rima oris is surrounded by upper and lower lips that come together laterally at the angle. At the caudal part of the tongue the oral cavity is continuous with the oropharynx.
The distal portion of the adenohypophysis expands by forming branching epithelial cords between which is very vascular mesenchymal tissue. Subsequent expansion causes the residual lumen to disappear. The wall of the adenohypophysis adjacent to the neurohypophysis remains thin and is called the intermediate portion. The stem remnant disappears.
The dental lamina invades the underlying mesenchyme and forms 10 bell shaped enamel organs in each jaw that represent the beginning of the deciduous (milk) teeth. The enamel organ subsequently becomes indented on its deep side by a mesenchymal condensation called the dental papilla. Each enamel organ and dental papilla together make up a tooth bud. The outer enamel layer of a tooth is produced by the enamel organ. The inner dentine layer and the cementum are formed by the dental papilla. The buds for the permanent teeth form in a similar manner during the third month on the lingual aspect of the deciduous teeth. The labiogingival band also invades the jaw mesenchyme and becomes a sulcus that separates the lip and cheek from the gingiva or gum area. The sulcus will enlarge to become the vestibule of the oral cavity.
The distal segment of the parotid bud begins to branch around the facial nerve as it grows toward the preauricular area. It represents the parotid gland. The proximal segment becomes the parotid duct that in the adult empties into the upper part of the vestibule at the upper second molar tooth. The submandibular bud develops in a similar manner in the floor of the oral cavity giving rise to the submandibular gland and duct. The sublingual gland sprouts from the proximal part of the bud near the oral epithelium.
B. FOREGUT
The caudal part of the lateral palatine processes will subsequently fuse to form the soft palate and uvula that separate the nasopharynx from the oropharynx. The auditory tube (pouch 1) opens into the lateral wall of the nasopharynx. The more caudal palatine fossa (pouch 2) is located in the lateral wall of the oropharynx. The fossa will be invaded at a later period by lymphocytes that produce the palatine tonsil.
The greater thymic bud (pouch 3) migrates into the cranial part of the thorax where it joins its counterpart from the opposite side to form the thymus. The thymus comes to rest between the sternum and the left brachiocephalic vein but may extend as far cranially as the thyroid gland. As the greater thymic bud migrates caudally, it pulls the caudal (inferior) parathyroid bud (pouch 3) to a lower position than the rostral (superior) parathyroid bud (pouch 4). Each parathyroid bud will become a parathyroid gland on the dorsal surface of the thyroid gland.
The laryngeal pharynx continues caudally as the esophagus at the level of the cricoid cartilage. The four layers (mucosa, submucosa, muscularis and fibrosa) of the esophagus become apparent. The esophagus lies dorsal to the trachea and to the right of the aorta. As it approaches the diaphragm, it courses to the left in front of the aorta.
Stomach rotation is complete with the greater curvature facing caudally and the lesser curvature facing cranially. The stomach can be divided into three portions, fundus body and pyloric end. The pylorus develops at the gastroduodenal junction.
The dorsal pancreas becomes the body of the pancreas, which passes to the left to end as the tail at the spleen. The body subsequently fuses with the ventral surface of the left kidney.
RESPIRATORY SYSTEM
The thyroid cartilage forms as a plate of cartilage joined in the midline with its counterpart on the opposite side. It lies in front of the laryngeal orifice. The arytenoid cartilage appears in the arytenoid swelling dorsal to the orifice. The laryngeal cartilages (thyroid, arytenoid, corniculate and cuneiform) are derived from the condensations representing the fourth and fifth arch cartilages. The vocal folds (cords) grow medially from the lateral wall of the larynx and are separated by the glottis. The infraglottic cavity lies caudal to the vocal folds surrounded by the cricoid cartilage. The cricoid cartilage is considered to represent a derivative of a sixth arch cartilage.
The trachea begins at the cricoid cartilage and its walls are composed of a series of well-defined, U shaped cartilages. The caudal movement of the tracheal bifurcation slows and later attains the definitive level of T-4–T-5 vertebrae.
The bronchopulmonary buds (tertiary branches) branch repeatedly within the lung sac giving rise to the lung containing many generations of progressively smaller intrapulmonary bronchi. Approximately 18 generations are produced by the time of birth, the very small ones being called bronchioles. Between the sixth month and term, the distal ends of the terminal bronchioli expand to form the alveoli where gaseous exchange with the blood will take place after birth.
C. MIDGUT
The massive liver lies beneath the diaphragm and occupies approximately half of the abdominal cavity. It is very vascular and is the major site where blood cells are produced (hematopoiesis). Before birth hematopoietic activity in the liver subsides and moves to the red bone marrow.
The ventral pancreas becomes the head of the pancreas that joins the body around the superior mesenteric vein. Later the duct system of the dorsal and ventral pancreas fuse. The main pancreatic duct is formed from the ventral pancreatic duct and the distal segment of the dorsal pancreatic duct. It empties into the duodenum with the bile duct at the major papilla. The proximal segment of the dorsal duct either disappears or persists as the narrower accessory pancreatic duct that empties into the duodenum at the minor papilla.
All of the jejunum and most of the ileum arise from the cranial limb of the midgut loop. The caudal limb becomes the terminal segment of the ileum, the cecum with its appendix, the ascending colon and the right two-thirds of the transverse colon. The midgut loop remains in the umbilical cord until the third month when it returns to the abdominal cavity. The cranial limb returns first with the jejunum settling on the left and the ileum on the right. The caudal limb then returns. The cecum, appendix, ascending colon and descending colon initially lie from right to left in a horizontal position below the liver. The cecum and appendix gradually descend from the upper right to the lower right, pulling the ascending colon with it. See Section VI for fixation of the midgut derivatives to the dorsal body wall.
D. HINDGUT
When the midgut loop returns to the abdominal cavity, the cranial part of the hindgut is pushed to the left. It gives rise to the left one-third of the transverse colon, the descending colon and the sigmoid. The colon is very narrow initially but later will dilate to form the large intestine.
The terminal segment of the rectum gives rise to the upper part of the anal canal, which is therefore lined with endoderm. The lower part of the anal canal is derived from the proctodeum and is lined with ectoderm. The two parts of the anal canal become continuous with the disappearance of the anal membrane. The anal canal opens to the exterior at the anus. In the adult the point of attachment of the anal membrane is represented by the pectinate line. The pectinate line is the viscero-somatic junction with mucosa above and specialized skin below. The innervation, blood supply and lymphatic drainage differ for the two adjacent areas.

FIG. 8 –2 [printable version of FIG 8-2]
- Alimentary and respiratory systems of the 30-mm embryo. The stomach is to the left and ventral to the duodenum and pancreas.
- Heart and great vessels of the 30-mm embryo.
- Urogenital system of the 30-mm embryo.
IV. UROGENITAL SYSTEM
Fig. 8–2CA. MESONEPHROS
The number of segments occupied by the mesonephros is further reduced, extending approximately from T-11 to L-1. The capsules and tubules at the cranial end disappear. Only the more caudal tubules persist and come to lie close to the lateral side of the gonad. See Section IV, E below for their fate in each sex.
Evidence that the human mesonephros functions as an excretory organ during the embryonic period has not been demonstrated.
B. KIDNEY (METANEPHROS)
The collecting tubules continue to divide at the periphery of the metanephros until approximately 12 generations are formed by the end of the fifth month. The second, third and fourth generations enlarge and coalesce to form the minor calyces of the renal pelvis. The tubules of the fifth generation become the collecting tubules that empty into the minor calyces.
Cell groups in the renal capsule differentiate into small spheres called renal vesicles in the vicinity of the collecting tubules. Each vesicle will form a nephron, which is the functional unit of the kidney. Each renal vesicle first becomes S shaped. The lower part of the S is invaginated subsequently by blood vessels forming a renal capsule (glomerulus and glomerular capsule). The upper part of the S will become a secretory tubule that establishes a communication with one of the collecting tubules. The secretory tubule will elongate and form the proximal convoluted tubule, the loop of Henle and the distal convoluted tubule.
The kidney is functional in the late fetal period.
C. BLADDER AND URETHRA (UROGENITAL SINUS)
The bladder portion of the urogenital sinus becomes the definitive bladder as the proximal segment of the ureter and mesonephric duct become incorporated into its lower wall to form the trigone. During the incorporation the orifice of the ureter moves cranial to the orifice of the mesonephric duct.
In the male the pelvic portion of the urogenital sinus will become the prostatic and membranous parts of the urethra. The phallic portion forms the penile urethra after the urogenital folds fuse on the underside of the phallus (penis). In the female the pelvic portion gives rise to part of the urethra and vagina. The phallic portion remains open thus forming most of the vaginal vestibule into which open the urethra and eventually the vagina.
By the end of the third month buds of the epithelial lining of the urethra extend into the surrounding mesenchyme. These buds will form the prostate gland in the male and the urethral and paraurethral glands in the female.
D. GONAD (DIFFERENT STAGE)
The gonad by its microscopic appearance, can be determined to be a testis in a genetically male embryo or an ovary in a genetically female embryo.
TESTIS
The gonadal cords elongate into discrete, solid testis cords that communicate with one another. The cords join a network of minute cords near the mesonephric remnant that will canalize at puberty and form the rete testis.
The testis cords are made up of germ cells and sustentacular cells (Sertoli). At puberty a lumen appears in the cords converting them into seminiferous tubules and the germ cells begin producing sperm. The seminiferous tubules empty into the rete testis, which in turn empties into the duct remnants of the mesonephros.
A tough capsule of dense connective tissue called the tunica albuginea begins to develop around the testis. Mesenchymal cells between the testis cords will differentiate into the interstitial cells (Leydig).
OVARY
The gonadal cords extend into the central part of the ovary where they break up into cell clusters containing groups of primordial germ cells. The cell clusters disappear and are replaced by a vascular stroma that forms the definitive ovarian medulla. No rete ovarii is formed.
The surface cells continue to proliferate and give rise to a second generation of cords called cortical cords. These cords remain close to the surface where they break up and surround the naked oogonia (germ cells). The surrounding cells will become the follicular cells.
E. GENITAL DUCTS
The mesonephric duct, mesonephric tubules and female duct develop differently in the two sexes. Differences in the pattern of development are apparent in the eighth week.
MALE
As the mesonephros regresses, five to 10 of the tubules adjacent to the testis make contact with the rete testis and become the efferent ducts. The efferent ducts drain into the mesonephric duct, which terminates in the upper (prostatic) part of the urethra and mainly becomes the ductus deferens (vas). The orifice of the mesonephric duct in the urethra moves closer to the midline. The segment of the mesonephric duct that receives the efferent ducts will become highly convoluted to form the ductus epididymis. Near its termination in the urethra an outpocketing will develop that produces the seminal gland (vesicle). Between the seminal gland and the urethra the mesonephric duct is called the ejaculatory duct. Caudal to the testis, vestiges of the mesonephric tubules will become the paradidymis.
The female duct regresses and most of it eventually disappears. Its cranial end will persist as the appendix testis. Its caudal end fuses with its counterpart on the opposite side. Some embryologists believe this portion is represented in the adult by a small diverticulum of the prostatic urethra called the prostatic utricle.
FEMALE
The mesonephric duct and tubules regress and will mainly disappear. Small remnants of the cranial portion may persist as the epoophoron and paroophoron in the mesentery of the ovary (mesovarium).
The female duct enlarges and canalizes. It can be divided into three segments: a) a cranial longitudinal segment that opens into the peritoneal cavity; b) an intermediate horizontal segment that passes toward the midline ventral to the mesonephric duct and c) a caudal longitudinal segment that fuses in the midline with its counterpart on the opposite side. The female ducts together assume a Y shape. The first two segments form the uterine tube, which opens into the peritoneal cavity at the abdominal ostium. As the intermediate segment passes medially, it brings with it a fold of tissue that will become the broad ligament of the uterus. The gonad lies dorsal to the fold or ligament. The fused caudal segment is the uterine primordium that will give rise to the uterus. Initially a septum passes through the middle of the primordium, which disappears thus forming the uterine cavity.
The caudal segment of the female duct makes contact with the dorsal wall of the pelvic portion of the urogenital sinus. The wall in the area of contact proliferates forming the sinus (Müllerian) tubercle. In the ninth week the tubercle gives rise to two solid evaginations called sinuvaginal bulbs. The bulbs fuse to form a solid vaginal plate with which the uterine primordium is continuous. A lumen begins to form in the plate by the end of the third month converting it into the vagina. By the end of the fifth month the vagina is entirely canalized. Its upper portion expands around the lower part of the uterus (cervix) to form fornices. A thin membrane called the hymen initially separates the vaginal canal from the vestibule of the vagina. It ruptures at a variable time after birth.
F. EXTERNAL GENITALIA
When the cloacal membrane disappears, the phallic part of the urogenital sinus opens to the exterior through the urethral groove and the rectum opens to the exterior through the anal canal and anus. Differences appear in the external genitalia (perineum), which permit sexual identification during the eighth week.
MALE
The genital tubercle elongates and enlarges into the phallus, which will become the penis. Mesenchymal tissue within the phallus organizes into two very vascular erectile masses called corpora cavernosa. Each corpus cavernosum attaches to the pubic bone by a crus. The urogenital groove and folds extend onto the underside of the phallus. The epithelial lining of the groove produces a urethral plate. By the end of the third month the urogenital folds fuse forming a raphe and converting the urethral plate into the penile part of the urethra. The penile part of the urethra is surrounded by erectile tissue called the corpus spongiosum that is continuous with the rounded glans penis at the distal tip of the phallus. During the fourth month an epithelial cord of cells at the tip of the glans penis grows deep to join the penile urethra. The cord soon canalizes whereby the penile urethra opens to the exterior through the meatus on the glans penis. The proximal part of the corpus spongiosum enlarges to form the bulb of the penis.
The labioscrotal swelling becomes the scrotal swelling. The swelling will fuse in the midline with its counterpart on the opposite side to form the scrotum into which the testes will later descend (see below). A scrotal septum will develop in the fusion area and separate the two testes. The fusion area is indicated on the surface by a raphe that joins with the raphe above and together they course from the anus to the glans penis.
FEMALE
The genital tubercle elongates and enlarges to a lesser extent producing a smaller phallus that will become the clitoris. The glans clitoris develops as a small rounded area at the distal tip of the phallus. Both the clitoris and its glans are also composed of erectile tissue. The urogenital folds do not fuse but remain open to become the labia minora. A mass of erectile tissue called the bulb of the vestibule develops at the base of each labium minus. The urogenital groove and phallic portion of the urogenital sinus together form the vestibule of the vagina located between the labia minora.
The labioscrotal swellings enlarge and remain separated thereby forming the labia majora.
G. DESCENT OF THE GONADS
By the end of the second month the gonads in both sexes begin to shift caudally in relation to the expanding abdominal wall. Since the initial blood supply from the aorta in the upper abdominal region is essentially retained, both the testicular and ovarian arteries descend from their original level to the testis and ovary, respectively.
The once broad urogenital mesentery, which attaches the mesonephros and gonad to the dorsal body wall, becomes very thin. Its cranial portion disappears along with the mesonephros. Its caudal portion becomes the gubernacular fold and extends caudally to the inguinal region. A band of mesenchyme runs from the caudal pole of the gonad, through the base of the gubernacular fold and inguinal region, into the labiogenital swelling. This band is called the gubernaculum, which is the "governor" of gonadal descent.
-
The change in position of the testis is greater than the ovary. The cause of the descent is not clear but it has been shown to be under hormonal influence.
Testis—By the end of the third month the testis lies behind the peritoneum near the inguinal part of the abdominal wall. A diverticulum of the peritoneal cavity called the vaginal sac (process) invades the gubernacular mesenchyme in the inguinal wall and marks the position of the inguinal canal. During the seventh month the testis passes through the canal but remains behind the peritoneum of the vaginal sac. When the vaginal sac reaches the scrotal swelling, it converts the swelling into a sac that receives the testis usually by the end of the eighth month. The lower part of the vaginal sac lines the scrotal cavity around the testis with a serous membrane called the tunica vaginalis testis. The upper part disappears when its walls come together thereby isolating the scrotal cavity from the peritoneal cavity. After the testis descends completely, the only remnant of the gubernaculum is a short, broad ligament between the caudal pole of the testis and the scrotal sac. It is necessary that the testis descends completely if it is to actively produce sperm at puberty.
Ovary—The ovary descends only as far as the brim of the true pelvis. Its gubernaculum becomes the ligament of the ovary and the round ligament of the uterus. The two ligaments together form a continuous band that runs from the caudal pole of the ovary to the labium majus.
V. COELOMIC CAVITY
The diaphragm domes over the expanding liver and attaches farther caudally on the dorsal wall than it does on the ventral wall. As a result the pleural cavity extends farther caudally at the dorsal wall between the ribs and the diaphragm with no intervening lung. This part of the pleural cavity is referred to as the costodiaphragmatic (pleural) recess.
As the urinary bladder and rectum enlarge near the caudal end of the peritoneal cavity, a peritoneal recess is produced between these viscera. In the male the recess is called the rectovesicle pouch (Douglas). In the female the recess is subdivided by the uterus and broad ligament into a ventral vesicouterine pouch and a dorsal rectouterine pouch.
VI. MESENTERIES
-
During the third month, after the herniated midgut derivatives return to the abdominal cavity from the umbilical cord, the dorsal mesentery fuses with the dorsal body wall in three areas. The three areas are as follows:
Most of the dorsal mesogastrium between its midline attachment and the spleen together with the mesoduodenum containing the pancreas swing to the left and come to lie adjacent to the left suprarenal gland and kidney. Since the suprarenal glands and kidneys are covered with peritoneum on only their ventral surface, they are said to be retroperitoneal in position. The segment above of the dorsal mesogastrium and mesoduodenum fuses with the ventral surface of the inferior vena cava, aorta, left suprarenal gland and left kidney. As a result most of the duodenum and pancreas become retroperitoneal. The short mesentery segment between the left kidney and spleen does not fuse thereby forming the lienorenal ligament. The tail of the pancreas and splenic blood vessels pass through this ligament to reach the hilum of the spleen. The segment of the dorsal mesogastrium between the stomach and the spleen is called the gastro-lienal ligament and contains gastric branches of the splenic artery. The lienorenal and gastrolienal ligaments form the left boundary of the lesser sac.
The mesentery of the midgut extends from the duodenal-jejunal junction to the distal segment of the transverse colon. It can be divided into a proximal, intermediate and distal portion. Only the small intermediate portion fuses with the dorsal body wall. The proximal portion suspends the jejunum and ileum. It becomes the mesentery proper that attaches obliquely to the dorsal body wall from upper left to lower right and contains most of the superior mesenteric vessels. As the cecum and ascending colon move into their definitive position at the far right, their mesentery and blood vessels fuse with the right side of the dorsal wall covering over the right ureter and part of the duodenum and right kidney. The distal portion suspends the transverse colon and persists as the transverse mesocolon. It attaches to the dorsal wall in a horizontal manner at the level of the pancreas and fuses with the dorsal aspect of the greater omentum.
The mesentery of the hindgut extends from the distal segment of the transverse colon to the rectum. It suspends all of the hindgut derivatives (distal segment of the transverse colon, descending colon and sigmoid) and contains the inferior mesenteric vessels. The hindgut derivatives are pushed to the far left side by the returning midgut. The mesentery of the descending colon and its contained vessels fuse to the left side of the dorsal wall covering over the left ureter and caudal part of the left kidney. The mesentery attached to the sigmoid persists as the sigmoid mesocolon.
VII. DIAPHRAGM
The diaphragm becomes dome shaped as it is pushed cranially into the pleural cavities by the expanding liver. It is attached at its periphery to the sternum, lower costal cartilages and vertebral column. Its attachment to the vertebral column is accomplished by way of two vertical muscular bands called right and left crura. Primordial muscle cells make up the diaphragm except in the area between the heart and liver called the central tendon.
VIII. CARDIOVASCULAR SYSTEM
A. HEART
Fig. 8–2BRIGHT ATRIUM
The cranial portion of the right venous valve (valve of sinus venosus) along with the septum spurium will form the crista terminalis in the wall of the adult right atrium. The crista terminalis extends ventrally along the right border between the orifices of the superior and inferior venae cavae and marks the boundary between the primitive atrium and sinus venosus. The primitive atrial portion of the right atrial wall is located ventral to the crista terminalis and contains musculi pectinati. The sinus venosus portion is located dorsally and has a smooth wall.
The caudal portion of the right venous valve fuses with the septum between the orifices of the inferior vena cava and coronary sinus.
The intermediate portion of the right venous valve becomes the valve of the inferior vena cava and valve of the coronary sinus. Highly oxygenated blood returning from the placenta is shunted by way of the valve of the inferior vena cava toward the foramen secundum to the left atrium.
LEFT ATRIUM
The common pulmonary vein is absorbed into the dorsal wall of the left atrium giving separate orifices for the right and left pulmonary veins. Subsequently the right and left pulmonary veins are also absorbed into the dorsal wall of the left atrium as far as their upper and lower tributaries. Their incorporation into the left atrial wall produces a total of four pulmonary vein orifices.
The primitive atrium with the musculi pectinati in its wall is greatly reduced on the left side becoming mainly the auricle of the left atrium. The absorbed pulmonary veins form most of the left atrial wall that is smooth and located dorsally.
INTERATRIAL SEPTUM
The septum secundum thickens. Its caudal, free edge becomes crescent shaped and is directed toward the orifice of the inferior vena cava, which also helps shunt the highly oxygenated blood from the placenta to the left atrium.
The lower free edge of the septum secundum eventually overlaps the foramen secundum converting it into an oblique opening bound by the septum secundum and septum primum. The interatrial opening becomes the foramen ovale that remains patent until birth. After the foramen is closed by the thin septum primum, the depression on the right side of the interatrial septum is called the fossa ovalis. The arching free edge of the septum secundum is the annulus ovalis.
B. ARTERIES
The majority of the definitive, named arteries are present by the end of the eighth week. Those in the extremities lag in attaining the adult pattern. The right and left coronary arteries are prominent, first branches of the aorta (ascending part). They arise immediately above the aortic semilunar valve. Each artery courses primarily in its respective part of the atrioventricular (A-V) sulcus sending major branches into the interventricular sulcus.
C. VEINS
VEINS IN THE LIVER
After the ductus venosus closes at birth, its afferent branches in the liver become branches of the portal vein thereby causing blood from the gut to flow to the hepatic sinusoids. The efferent veins that drain the sinusoids become tributaries of the hepatic veins that drain into the inferior vena cava.
DURAL SINUSES
As the otic capsule expands, the sinuses dorsal to the capsule enlarge and connect the tributaries of the middle and posterior cerebral plexuses through the primitive transverse and sigmoid sinuses. The anterior cerebral plexus drains predominantly into the middle cerebral plexus. That portion of the primary head vein medial to the trigeminal ganglion becomes the definitive cavernous sinus, which establishes new connections with the primitive sigmoid sinus known as the superior and inferior petrosal sinuses. The midline portions of the cerebral plexuses enlarge to produce new sinuses (superior sagittal, inferior sagittal and straight) that connect with the primitive transverse sinus. The primitive sinuses move with the expanding cerebral hemispheres attaining the definitive position together.
PULMONARY VEINS
The right and left pulmonary veins disappear as they become incorporated into the left atrial wall. Their upper and lower tributaries become the upper and lower right and upper and lower left pulmonary veins.
LYMPHATICS
The mesenteric lymph sac in the upper lumbar segments will dilate to form the definitive cisterna chyli. The left jugular lymph sac and cisterna chyli grow toward one another to form the major lymph channel of the body, the thoracic duct.
CIRCULATORY CHANGES AT BIRTH
Two important events occur at birth that profoundly influence the newborn’s circulatory system: a) the flow in the umbilical vessels is interrupted when the placenta breaks away from the uterine wall and b) the newborn begins breathing thereby opening the capillary lung field, which greatly increases the pulmonary circulation. The physiological changes are rapid, occurring within a few days. The anatomical changes usually take place over several months.
When the blood flow through the lungs increases, the pulmonary venous return to the heart increases thereby raising the pressure in the left atrium. This presses the thin septum primum against the thick septum secundum and functionally closes the foramen ovale. Anatomical closure produces the fossa ovalis. Simultaneously the muscular wall of the ductus arteriosus contracts increasing the flow to the lungs through the pulmonary arteries. Within three months its lumen is obliterated thereby producing the ligamentum arteriosum.
The muscular wall of the umbilical arteries contracts when blood flow through them ceases. Anatomical closure converts their distal segment on the deep side of the ventral abdominal wall into the medial umbilical ligaments. Their proximal segment gives rise to the superior vesical arteries to the bladder. The umbilical vein and ductus venosus close a short time after the umbilical arteries. Anatomical closure changes the umbilical vein into the ligamentum teres that courses from the deep side of the umbilicus, through the edge of the falciform ligament, to the portal vein where the latter enters the liver (hilum). The ductus venosus becomes the ligamentum venosum between the portal vein and the inferior vena cava.
IX. SKELETAL SYSTEM
A. SKULL
NEUROCRANIUM
Cartilaginous Neurocranium
The parachordal plate becomes the basioccipital on the ventral aspect of the medulla oblongata. The hypoglossal nerve (XII) emerges from the skull through the hypoglossal canal in its caudolateral portion that arises from the occipital sclerotomes. These sclerotomes extend lateral to the hypoglossal canal as the exoccipital that forms the lateral boundary of the foramen magnum at the brain-spinal cord junction. The exoccipital blends with the supraoccipital that closes the foramen magnum dorsally. Most of the definitive occipital bone will arise from the basi-, ex- and supra-occipital.
The basi- and exoccipital join the otic capsule laterally, which will become the petrous part of the temporal bone in the adult. Cranial nerves IX and X and the spinal accessory nerve leave the skull along with the internal jugular vein through the jugular foramen between the exoccipital and otic capsule. Cranial nerves VII and VIII leave the cranial cavity through the internal auditory meatus between the basioccipital and otic capsule. The petrous part of the temporal bone houses the entire membranous labyrinth. The endolymphatic duct will emerge from the bone through a small slit near the fourth brain ventricle.
The hypophyseal cartilages fuse around the hypophysis to form the body of the sphenoid bone containing the hypophyseal fossa (sella turcica). The rostral end of the basioccipital joins with the body of the sphenoid bone. The alisphenoid will become the greater wing of the sphenoid bone through which the maxillary (V2) and mandibular (V3) nerves pass from the skull by way of the foramen rotundum and foramen ovale, respectively. The alisphenoid joins the capsule as the alicochlear commissure that passes lateral to the internal carotid artery. The orbitosphenoid expands as a plate supporting part of the cerebral hemispheres. It will become the lesser wing of the sphenoid bone containing the optic canal through which the optic nerve (II) passes to the eye. The oculomotor (III), trochlear (IV), ophthalamic (V1) and abducens (VI) nerves will enter the orbit through the superior orbital fissure between the alisphenoid and orbitosphenoid.
The trabecular cartilage and nasal capsule will give rise to the ethmoid bone and most of the nasal septal cartilage. The fascicles of the olfactory nerve (I) course through the cribriform plate of the ethmoid to the olfactory mucosa in the roof of the nasal cavity.
Membranous Neurocranium
The outer part of the ectomeninx becomes an osteogenic membrane that covers the dorsal and lateral surfaces of the brain. The osteogenic membrane grows as the cerebral hemispheres enlarge. It gives rise to the frontal, parietal, squamous temporal and interparietal occipital bones of the adult. These bones are separated by thin planes of connective tissue called sutures. At birth a wide suture area called the anterior fontanelle is located in the midline at the junction of the frontal and parietal bones. The anterior fontanelle disappears during the first few years of life.
VISCEROCRANIUM
The malleus and incus ear ossicles develop in the dorsal segment of the first arch cartilage. The anterior ligament of the malleus and sphenomandibular ligament are adult remnants of its intermediate segment. The mandible is laid down lateral to the ventral segment, which eventually disappears.
The stapes ear ossicle develops in the dorsal segment of the second arch cartilage. The ear ossicles are the first bones to fully ossify and reach their adult size. This occurs around the fourth month. The styloid process and stylohyoid ligament are remnants of its intermediate segment. The lesser horn and body (cranial part) of the hyoid bone develop from its ventral segment after it fuses in the midline with its counterpart on the opposite side.
The third arch cartilage also fuses in the midline with its counterpart on the opposite side and with the median part of the fused second arch cartilages. It gives rise to the greater horn and body (caudal part) of the hyoid bone.
The fourth, fifth and sixth arch cartilages never become distinct cartilaginous bars. Instead the laryngeal condensations that represent them give rise directly to the laryngeal cartilages. The thyroid, arytenoid, corniculate and cuneiform are thought to be derived from the fourth and fifth cartilages. The cricoid supposedly represents the sixth cartilage.
B. POSTCRANIAL PART OF AXIAL SKELETON
VERTEBRAL COLUMN
The notochord degenerates in the area of the centrum but expands in the intervertebral disc to form the nucleus pulposus.
The neural arch fuses with its counterpart on the opposite side dorsal to the spinal cord. Together they produce a spinous process flanked by a lamina on each vertebra.
The S-1 to S-5 sclerotome segments fuse into one bone, the sacrum. The first segment extends laterally as a wing shaped process called the ala. The lateral border of the sacrum articulates with the pelvis at the sacroiliac joint.
RIBS AND STERNUM
The long, proximal segment of each rib primordium begins ossifying to produce a rib. The short distal segment remains cartilaginous forming a costal cartilage. All but the last two costal cartilages attach directly or indirectly to the sternum.
The sternal bar and episternal cartilage will eventually fuse with their counterpart on the opposite side to produce the body and manubrium of the sternum, respectively. Initially the sternum is cranial to its definitive level because of the large liver preventing its caudal displacement.
C. APPENDICULAR SKELETON
The clavicle becomes a prominent, well-ossified bone that attaches the scapula (acromion process) to the manubrium of the sternum.
The spine of the scapula becomes recognizable. The head of the humerus articulates with the scapula at the glenoid fossa.
The pelvic bone is represented by three pairs of cartilages: ilium, ischium and pubis. The pubic cartilages are separated in the midline by the symphysis pubis. All of the cartilages enter into the formation of the acetabular fossa where the head of the femur articulates.
Most of the bones of the extremities can be identified and are mainly cartilagenous. Before birth the majority of the primary ossification centers occur. The bones of the upper extremity are humerus (arm), radius and ulna (forearm), carpals (wrist), metacarpals (palm) and phalanges (fingers). The bones of the lower extremity are femur (thigh), patella (knee), tibia and fibula (leg), tarsals (ankle), metatarsals (sole) and phalanges (toes).
X. MUSCULAR SYSTEM
All of the definitive skeletal muscles and their innervation can be identified by the end of the eighth week. Stimulation studies have shown that some of the skeletal muscles are reflexly active at this time. Motor nerve fibers are in the vicinity of the premuscle cells at the time the premuscle cells begin to differentiate. Most contractions of premuscle cells are probably neurogenic in origin.
The muscles of mastication (masseter, temporalis, medial pterygoid and lateral pterygoid) differentiate from the premuscle mass in branchial arch 1. The premuscle cell laminae and facial nerve branches from branchial arch 2 spread into the face from the lateral aspect of the head. Facial muscles begin to differentiate laterally in the lower face, gradually proceeding to the midline and to the upper face. Around the end of the third month all of the definitive facial muscles can be identified in their definitive position.
XI. INTEGUMENTARY SYSTEM
A. SKIN
Definitive skin is divided into an outer layer, the epidermis, and the adjacent deep layer, the dermis. The epidermis is derived from the single cell layer of ectoderm on the surface. By the end of the second month the ectoderm (basal layer) produces at the surface a layer of flattened cells called the periderm. During the third month the basal layer also forms an intermediate layer. Four definitive layers of the epidermis can be identified by the fourth month and are produced by the basal and intermediate layers. From deep to superficial they are the stratum basal, stratum spinosum, stratum granulosum and stratum corneum. Neural crest cells are thought to invade the epidermis during the third month and give rise to the melanoblasts that produce melanin (skin pigment). The periderm is shed during the second half of pregnancy when the epidermis produces a protective, white, cheesy substance called vernix caseosa
The dermis (corium) develops from the underlying mesenchymal tissue that is probably derived for the most part from dermatomes. Connective tissue fibers differentiate in the mesenchymal tissue during the third and fourth month. The superficial layer or stratum papillare extends into the epidermis as dermal papillae that contain capillaries and sensory receptors. The deep layer, stratum reticulare, is composed primarily of connective tissue in which fat accumulates before term.
B. NAILS
Each nail on the fingers and toes begins during the third month as an epidermal thickening at the tip of the digit called the nail field. As the digits lengthen, the nail field migrates to the dorsal side bringing along the palmar or plantar nerves. An epithelial nail fold develops at the periphery of the nail field and extends proximally beneath the epidermis to form the nail matrix. During the fifth month the nail matrix produces special, keratinized cells that compact to form the nail plate. As the nail plate is produced, it moves distally over the nail bed reaching the tip of the digit by term.
C. HAIR
Fine fetal hair (lanugo) begins to develop during the early part of the third month beginning in the face. By the end of the third month it is scattered over much of the body. Fetal hair is shed around term and is replaced by coarser infant hair.
Each hair begins as a bud or cylindrical downgrowth of the basal layer of the epidermis into the underlying dermis. Each bud forms a hair follicle, the growing tip of which is invaginated by mesenchymal tissue. The mesenchyme produces a papilla containing capillaries and nerve terminals. The cells in the center of the follicle become keratinized forming the hair shaft. The peripheral cells give rise to the epidermal sheaths. Sebaceous glands arise as outpocketings of the peripheral follicle cells. The cells in the center of such outpocketing break down to produce sebum that is secreted into the follicle.
Sweat glands begin similar to hair but their downgrowth is not invaginated with mesenchymal tissue. Later they become coiled and canalized.
D. MAMMARY GLAND
During the fetal period (third month to term) the mammary gland primordium (primary bud) in both sexes penetrates the underlying tissue where it gives rise to 16 to 24 sprouts. Near term each sprout canalizes and becomes a lactiferous duct. Tertiary sprouts arise from the lactiferous duct and invade the underlying fat to become the ducts and alveoli of the gland.
Initially the lactiferous ducts open onto the surface into a depression called the mammary pit. After birth the surrounding tissue proliferates to produce the nipple.
E. TEETH
The labiodental lamina produces two additional laminae: an outer labial lamina that invades the maxillary and mandibular processes to form the labiogingival groove and an inner dental lamina that gives rise to buds called enamel organs. The labiogingival groove will enlarge to become the vestibule of the mouth between the lips and the jaws. Ten enamel organs are produced in each jaw and represent the primordia of the deciduous teeth.
During the third month the deep surface of each enamel organ is invaginated by the underlying mesenchyme. As a result of the invagination, the enamel organ becomes bell shaped and differentiates into three layers, an outer and an inner enamel epithelium separated by the enamel reticulum. The invaginated mesenchyme proliferates to form a condensed mass called the dental papilla.
The inner enamel epithelium gives rise to ameloblasts that produce enamel. After the tooth erupts, the enamel organ is shed. The cells of the dental papilla adjacent to the inner enamel epithelium become odontoblasts that form dentine deep to the enamel. The odontoblasts persist throughout the life of the tooth. Blood vessels and nerves invade the remaining part of the dental papilla forming the pulp.
By the end of the third month the enamel organs for the permanent teeth begin to develop on the lingual side of the deciduous teeth. The deciduous teeth start to erupt approximately 6 months after birth; the permanent teeth begin to erupt approximately 6 years after birth.
XII. SPECIAL SENSE ORGANS
A. EYE
OPTIC STALK
The nerve fibers coursing through the optic stalk increase in number and eventually completely obliterate its lumen thereby converting it into the optic nerve. The hyaloid artery passing through the stalk will give rise to the central artery of the retina.
The same protective membranes that form around the brain (viz. dura mater, arachnoid and pia mater) also develop around the optic nerve. The subarachnoid space containing cerebrospinal fluid extends peripherally to the retina around the optic nerve.
LENS
Secondary lens fibers are continually added to the nucleus of the lens throughout prenatal and much of postnatal life.
CAPSULAR MESENCHYME
Two layers begin to differentiate in the capsular mesenchyme: a) an inner, thin, vascular, pigmented layer called the choroid, and b) an outer, thick, white fibrous layer called the sclera.
ACCESSORY STRUCTURES
The upper and lower eyelid folds become thin, grow toward each other and begin to fuse. When fusion is complete (ninth week), a space is located between the surface of the eye and the eyelids called the conjunctival sac. The eyelids remain fused until about the seventh month of prenatal life.
The nasolacrimal duct courses from the medial aspect of the conjunctival sac to the nasal cavity. Its orifice in the nasal cavity is located below the inferior nasal concha in a space called the inferior meatus.
Small, solid epithelial buds begin to sprout from the upper, lateral part of the conjunctival sac. These buds are the lacrimal gland primordium and will increase in number, branch and canalize to form the ducts and ductules of the lacrimal gland. The lacrimal gland secretes fluid into the conjunctival sac for protection of the eye surface. Excessive fluid production is channeled to the nasal cavity by way of the nasocrimal duct.
B. EAR
INTERNAL EAR
The membranous labyrinth attains, for the most part, its definitive configuration. Two minor changes occur subsequent to the eighth week: a) the formation of the Y-shaped connection between the endolymphatic duct, utricle and saccule and b) the elongation and narrowing of the communication between the saccule and cochlear duct forming the ductus reunions.
Vestibular Pouch
One end of each semicircular canal enlarges to produce an ampulla at its junction with the utricle. A crest or crista will differentiate later within each ampulla that functions as a receptor for the maintenance of equilibrium. Similar receptor areas called maculae will also form in the walls of the utricle and saccule. Nerve fibers from both types of receptors have their cell bodies in the vestibular ganglion.
Cochlear Pouch
The cochlear duct completes the two and one half turn spiral (definitive number of turns). The receptor area for sound (spiral organ of Corti) will differentiate later from a ridge of modified epithelial cells in the cochlear duct. As the cochlear duct spirals, so follows the cochlear ganglion thereby producing the spiral ganglion. Nerve fibers from the receptor area for sound have their cell bodies in the spiral ganglion.
Otic Capsule
The membranous labyrinth is encased in cartilage. Perilymphatic spaces containing fluid develop between the membranous labyrinth and the cartilage. The cartilage will subsequently transform into bone forming the bony labyrinth.
MIDDLE EAR
The mesenchymal condensations representing the middle ear ossicles transform into cartilage thereby producing a physical connection between the tympanic membrane (see below) and the perilymphatic space around the cochlear duct.
The endodermal lining of the primitive tympanic cavity expands subsequently to form the definitive tympanic cavity. As it expands, it envelops the ossicles and chorda tympani nerve. Expansion dorsally produces the tympanic antrum and mastoid air cells.
EXTERNAL EAR
The auricular cartilages appear as cellular condensations when most of the definitive external features of the auricle become evident.
Epithelial cells lining the external acoustic meatus begin to form a meatal plug that eventually occludes the entire meatus. The plug dissolves around the seventh month of prenatal life.
The tympanic membrane separates the external acoustic meatus from the tympanic cavity. It is composed of three layers, ectoderm on the exterior, endoderm on the interior and an intermediate layer of mesenchymal cells in which the handle of the malleus is enmeshed.
C. OLFACTORY APPARATUS
THE 30-MM EMBRYO
STAGE 23
AGE: 52 TO 56.5 DAYS
CROWN TO RUMP LENGTH 30.7 MM
Carnegie collection 4570
Reference
Streeter GL: Developmental horizons in human embryos. Description of age groups XIX, XX, XXI, XXII and XXIII. Contrib Embryol Carnegie Instn 34:165–196, 1951
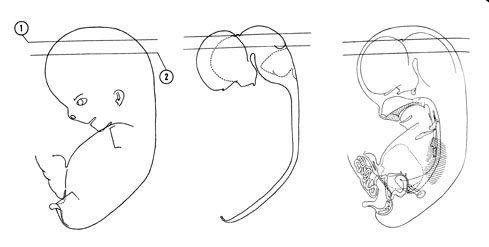
FIG. 8–3
SECTION 1
A section through the cerebral hemispheres (occipital lobe area), the roof of the diencephalon and the tectum of the mesencephalon.
Observe:
- The three zones of the cerebral hemisphere (neopallial cortex) and the tectum (collicular ridge) of the mesencephalon.
- Some of the components of the epithalamus of the diencephalon.
- The venous sinuses and plexus between the dura mater and the osteogenic layer.
- The small arteries coursing in the pia mater adjacent to the surface of the mesencephalon.
- The arachnoid trabeculae in the subarachnoid space where the cerebrospinal fluid circulates.
A section through the cerebral hemisphere (parietal and temporal lobe areas), dorsal thalamus and basis pedunculi area, tegmentum and tectum of the mesencephalon.
Observe:
- The choroid plexus in the lateral ventricle.
- Two areas of the third ventricle in the midline at the level of the diencephalon.
- The narrowed segment of the mesocoele where the cerebral aqueduct begins to form.
- The caudal part of the collicular ridge in the tectum on each side of the cerebellar vermis.
- The primordial nucleus of cranial nerve III in the tegmentum.
, the roof of the diencephalon and the tectum of the mesencephalon. A section through the cerebral hemisphere (parietal and temporal lobe areas), dorsal thalamus and basis pedunculi area, tegmentum and tectum of the mesencephalon.")
FIG. 8–3 [printable version of FIG 8-3]
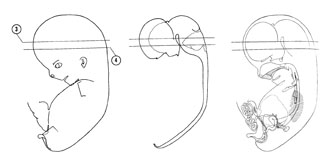
FIG. 8–4
SECTION 3
A section through the cerebral hemispheres (insular area and parietal and temporal lobe areas) and middle of the diencephalon and metencephalon.
Observe:
- The striatal ridge in the floor of the lateral ventricle and the primordial hippocampus in the medial wall of the cerebral hemisphere.
- The extension of the inferior horn of the lateral ventricle into the temporal lobe area.
- The hypothalamic sulcus demarcating the thalamus from the hypothalamus.
- The fusion area between the cerebral hemispheres and diencephalon.
- The intra- and extraventricular parts of the cerebellum.
A section through the cerebral hemispheres, hypothalamus and metencephalon.
Observe:
- The medial and lateral striatal ridges of the striatal part of the cerebral hemispheres.
- The interventricular foramen connecting the lateral and third ventricles.
- The amygdala area in the rostral part of the temporal lobe area.
- The primordial nuclei in the metencephalon.
- The choroid plexus and median aperture of the ependymal diverticulum in the roof of the fourth ventricle.
 and middle of the diencephalon and metencephalon. A section through the cerebral hemispheres, hypothalamus and metencephalon.")
FIG. 8–4 [printable version of FIG 8-4]
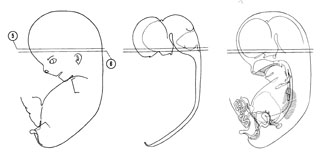
FIG. 8–5
SECTION 5
A section through the cerebral hemispheres, hypothalamus, metencephalon and medulla oblongata.
Observe:
- The rostral part of the medial and lateral striatal ridges.
- The large arteries in the subarachnoid space.
- The trigeminal root lateral to the pons area.
- The tract areas and primordial nuclei.
- The anterior semicircular canal in the otic capsule.
A section through the cerebral hemispheres (frontal lobe area), hypothalamus and medulla oblongata.
Observe:
- The lamina terminalis and preoptic area and recess of the third ventricle.
- The rostral part of the hypothalamus at the level of the optic chiasma.
- The neurohypophysis flanked on each side by the tuberal part of the adenohypophysis.
- The relation of cranial nerves III, IV, V, VII, VIII, IX, X and XI.
- Parts of the internal ear.
, hypothalamus and medulla oblongata.")
FIG. 8–5 [printable version of FIG 8-5]
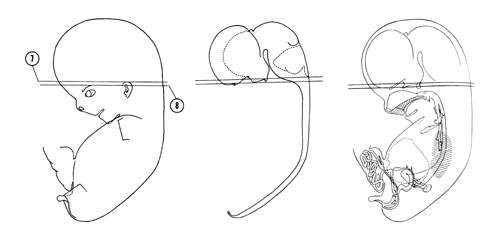
FIG. 8–6
SECTION 7
A section through the cerebral hemispheres, optic nerve, optic chiasma and medulla oblongata.
Observe:
- The adenohypophysis caudal to the optic chiasma and medial to the internal carotid artery and trigeminal ganglion.
- The relative position, parts and relation of the cochlear and vestibular portions of the internal ear.
- The trigeminal, geniculate and vestibular ganglia.
- The primordial nuclei and tract areas in the medulla oblongata where the fourth ventricle is continuous with the central canal.
- The roof of the orbit and its relation to the orbitosphenoid bone.
A section through the edge of the cerebral hemispheres (frontal lobe area) and medulla oblongata.
Observe:
- The olfactory bulb on the underside of the frontal lobe.
- The lacrimal gland primordium, ciliary ganglion, optic nerve and the lateral rectus muscle.
- The middle ear ossicles medial to the auricle.
- The cartilaginous components of the skull base.
- The primordial nuclei and tract areas in the medulla oblongata.
 and medulla oblongata.")
FIG. 8–6 [printable version of FIG 8-6]
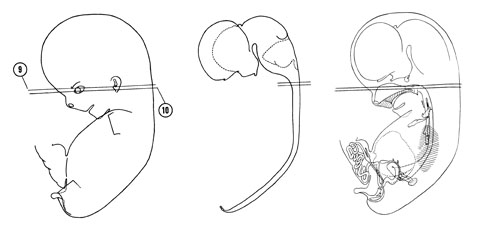
FIG. 8–7
SECTION 9
A section through the medulla oblongata and the cranial edge of the nasal cavity and nasopharynx.
Observe:
- The eyelid folds, conjunctival sac and two parts of the inner layer of the optic cup.
- The pterygopalatine and otic ganglia.
- The external acoustic meatus surrounded by the auricle and terminating at the tympanic membrane.
- The junction of the vertebral arteries forming the basilar artery on the ventral aspect of the medulla oblongata.
- The origin of the internal jugular vein at the jugular foramen and its relation to cranial nerves IX, X and XI.
A section through the middle of the eye and caudal part of the medulla oblongata.
Observe:
- The layers of the eye.
- The muscles of mastication between the eye and the auditory tube (pouch 1).
- The choana through which the nasal cavity communicates with the nasopharynx.
- The relation of the first and second arch cartilages to the middle ear.
- The hypoglossal nerve in the subarachnoid space.

FIG. 8–7 [printable version of FIG 8-7]
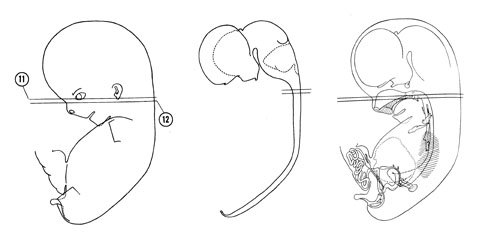
FIG. 8–8
SECTION 11
A section through the caudal part of the secondary palate, junction of the medulla oblongata and the spinal cord and the C-1 spinal ganglion.
Observe:
- The lower part of the eye.
- The dorsal part of the mandible and its relation to the first arch cartilage and parotid gland.
- The unfused uvula and soft palate parts of the secondary palate.
- The relations of the nerves, ganglia, blood vessels and muscles at the base of the skull.
- The C-1 spinal ganglion which usually disappears in the adult.
A section through the dorsal edge of the tongue and the C-2 spinal ganglion.
Observe:
- The vomeronasal organ on the medial wall and the inferior concha on the lateral wall of the nasal cavity.
- The relative positions of the nasal cavity, oral cavity, oropharynx and laryngeal pharynx.
- The palatine fossa (pouch 2) in the lateral wall of the oropharynx.
- An enamel organ and the maxilla in the roof of the oral cavity.
- The atlantoaxial joint.

FIG. 8–8 [printable version of FIG 8-8]
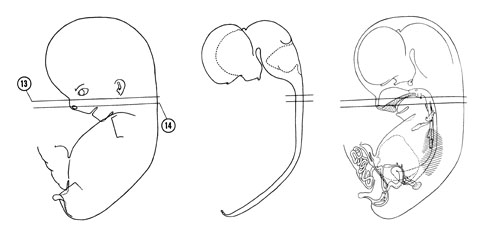
FIG. 8–9
SECTION 13
A section through the rostral part of the nasal cavity and secondary palate.
Observe:
- The fusion lines between the lateral palatine process and nasal septum.
- The well-developed genioglossus muscle in the tongue.
- The facial muscles and parotid duct beneath the ectoderm.
- The superior sympathetic ganglion, inferior vagal ganglion and internal and external carotid arteries between the sternocleidomastoid muscle and the pharynx.
- The junction of the jugular lymph sac and the internal jugular vein.
A section through the external nostrils, premaxillary palate, larynx and C-3 spinal ganglion.
Observe:
- Epithelial plugs in the external nostrils.
- The well-developed tongue muscles and the hypoglossal nerve.
- The submandibular duct, gland and ganglion.
- The arytenoid and thyroid cartilages of the larynx and the greater horn and body of the hyoid apparatus.
- The dorsal and ventral primary rami of a spinal nerve.

FIG. 8–9 [printable version of FIG 8-9]
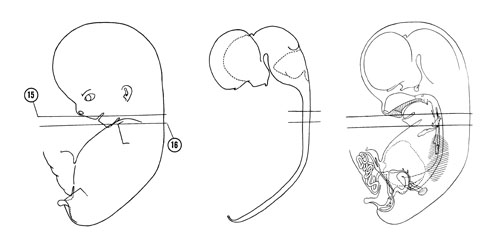
FIG. 8–10
SECTION 15
A section through the angle of the mouth, tongue tip and beginning of the esophagus.
Observe:
- The frenulum of the tongue lateral to which is the orifice of the submandibular duct.
- The lower facial muscles beneath the developing skin.
- The strap muscles of the neck ventral to the thyroid cartilage.
- The rostral parathyroid gland (pouch 4) dorsal to the thyroid gland.
- The cricoid cartilage surrounding the infraglottic cavity of the larynx.
A section through the edge of the chin, manubrium of the sternum and C-5 spinal ganglion.
Observe:
- The tips of the first arch cartilage in the edge of the chin.
- The clavicle connecting the acromion of the scapula with the manubrium of the sternum.
- The head of the humerus surrounded by the deltoid muscle.
- The isthmus of the thyroid gland ventral to the trachea and the caudal parathyroid gland (pouch 3) lateral to the trachea.
- The recurrent laryngeal nerve (motor to the laryngeal muscles) in the groove between the trachea and esophagus.

FIG. 8–10 [printable version of FIG 8-10]
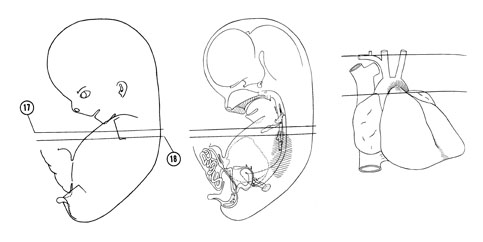
FIG. 8–11
SECTION 17
A section through the sternum, viscera at the root of the neck and C-7 spinal ganglion.
Observe:
- The bilateral thymus gland (pouches 3 and 4) deep to the sternum.
- The brachiocephalic veins about to join to form the superior vena cava.
- The bifurcation of the brachiocephalic artery next to the trachea into the right subclavian and right common carotid arteries.
- The origin of the vertebral artery from the right subclavian artery ventral to which the right vagus nerve courses.
- The left vagus nerve coursing between the left brachiocephalic vein and left common carotid artery.
A section through the caudal part of the sternum, cranial part of the heart and lungs and T-1 spinal ganglion.
Observe:
- The pulmonary semilunar valve separating the right ventricle and pulmonary trunk.
- The aortic arch passing obliquely to the left of the trachea and esophagus.
- The left vagus nerve coursing between the aortic arch and the left lung with its recurrent laryngeal branch looping around the arch and ascending to the larynx.
- The origin of the left subclavian artery from the aortic arch.
- The subdivisions of the grey and white matter in the spinal cord.

FIG. 8–11 [printable version of FIG 8-11]
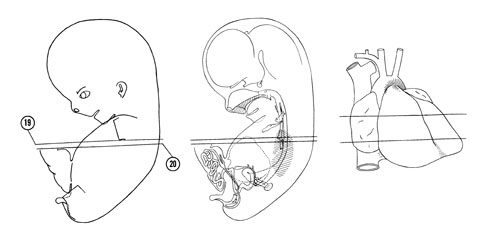
FIG. 8–12
SECTION 19
A section through the cranial edge of the liver and diaphragm, tracheal bifurcation and T-2 spinal ganglion.
Observe:
- The interventricular septum separating the right and left ventricular chambers.
- The aortic semilunar valve between the left atrium and ascending aorta where the coronary arteries arise.
- The junction of the superior vena cava in Section 18 with the sinous venosus part of the right atrium.
- The bifurcation of the pulmonary trunk into right and left pulmonary arteries.
- The ductus arteriosus connecting the pulmonary trunk with the aorta.
A section through the apex of the heart, root of the lungs and T-2–T-3 interganglion region.
Observe:
- The central tendon of the diaphragm between the heart and liver to which the pericardial sac attaches.
- The mitral valve between the left atrial and ventricular chambers.
- The left phrenic nerve coursing in the pericardial sac to the diaphragm.
- The oblique and horizontal fissures in the right lung dividing it into upper, middle and lower lobes.
- The three intercostal muscles in an intercostal space.

FIG. 8–12 [printable version of FIG 8-12]
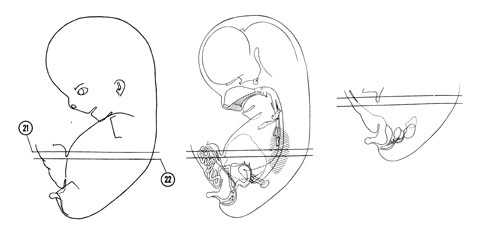
FIG. 8–13
SECTION 21
A section through the umbilical cord, caudal edge of the heart and the T-4 spinal ganglion.
Observe:
- The blood vessels, allantois and herniated gut in the umbilical cord.
- The right phrenic nerve to the right of the inferior vena cava at the diaphragm.
- The esophageal plexus formed by the vagus nerves.
- The oblique fissure in the left lung separating the upper lobe with its lingula from the lower lobe.
- The parietal and visceral subdivisions of the pleura lining the pleural cavity.
A section through the junction of the umbilical cord and the body wall and the T-6 spinal ganglion.
Observe:
- The near junction of the umbilical vein with the ductus venosus.
- An afferent vein of the liver arising from the ductus venosus, and efferent veins draining into the hepatic veins that, in turn, drain into the inferior vena cava.
- The reorganization of the esophageal plexus into anterior and posterior vagal trunks.
- The esophagus passing to the left, ventral to the aorta.
- The relation of the sympathetic trunk to the vertebral column and head of the ribs.

FIG. 8–13 [printable version of FIG 8-13]
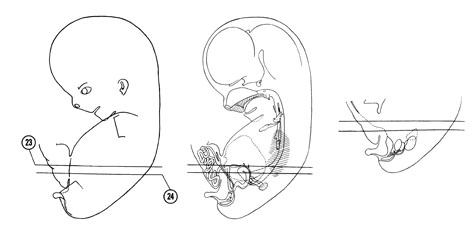
FIG. 8–14
SECTION 23
A section through the gall bladder, pylorus and T-7 spinal ganglion.
Observe:
- The jejunum passing into the umbilical cord and the colon passing back into the abdominal cavity.
- The pylorus and gastroduodenal junction where the pyloric sphincter develops.
- The hepatic ducts that drain bile from the liver and the cystic duct that drains the gall bladder.
- An afferent vein arising from the portal vein as it enters the liver carrying blood from the gut.
- The azygos vein on the ventral surface of the vertebral column.
A section through the esophageal-gastric junction, base of lungs and T-7–T-8 interganglion region.
Observe:
- The superior mesenteric artery passing between the jejunum and colon to the herniated midgut.
- The umbilical arteries entering the umbilical cord.
- The parts of the stomach; fundus, body and pyloric portion.
- The intrahepatic part of the inferior vena cava.
- The cranial edge of the suprarenal glands (cortex).

FIG. 8–14 [printable version of FIG 8-14]
FIG. 8–15
SECTION 25
A section through the pancreas, caudal edge of the lung and T-8 spinal ganglion.
Observe:
- The allantois entering the umbilical cord.
- The bile duct, portal vein and hepatic artery coursing dorsal to the head of the pancreas.
- The inferior vena cava between the liver and right suprarenal gland.
- The costodiaphragmatic recess in the caudal part of the pleural cavity.
- The muscles in the body wall.
A section through the celiac and T-9 spinal ganglia.
Observe:
- The obliterated segment of the allantois or urachus between the umbilical arteries.
- The superior mesenteric artery and vein coursing dorsal to the pancreas between its head and body.
- The celiac artery surrounded by the celiac ganglion.
- The cortex and medulla of the suprarenal gland and the close relation between the medulla and the celiac ganglion.
- The diaphragmic crus or muscle band on each side of the aorta as it enters the abdominal cavity.

FIG. 8–15 [printable version of FIG 8-15]
FIG. 8–16
SECTION 27
A section through the duodenal-jejunal junction, spleen and T-9–T-10 interganglion region.
Observe:
- The apex of the bladder that is continuous with the urachus in Section 26.
- The splenic vein passing dorsal to the body of the pancreas and the tail of the pancreas at the hilum of the spleen.
- The lesser sac enclosed by the greater omentum.
- The fusion area between the body of the pancreas and the left kidney and suprarenal gland.
- The superior mesenteric artery near its origin from the aorta.
A section through the cranial edge of the gonads, renal hilum and T-10 spinal ganglion.
Observe:
- The bladder flanked on each side by an umbilical artery.
- The caudal edge of the lesser sac.
- The left gonad covered by the greater omentum.
- The left renal vein between the duodenum and aorta coursing to the inferior vena cava.
- The superior mesenteric ganglion at the origin of the superior mesenteric artery from the aorta.

FIG. 8–16 [printable version of FIG 8-16]
FIG. 8–17
SECTION 29
A section through the caudal edge of the umbilical cord, liver and duodenum and the T-11 spinal ganglion.
Observe:
- The cut edge of the thigh on each side of the infraumbilical area of the abdominal wall.
- The arrangement of the muscles and aponeuroses in the abdominal wall.
- The origin of the right renal artery from the aorta.
- The major and minor calyces of the renal pelvis.
- The ganglion cells ventral to the aorta between the kidneys.
A section through the caudal part of the ventral abdominal wall, middle of the testis and T-11 spinal ganglion.
Observe:
- The junction of the thigh with the ventral abdominal wall.
- Some of the components of the testis and remnants of the mesonephros.
- The collecting tubules and renal corpuscles of the kidney.
- The origin of the ureter from the renal pelvis.
- The diaphragm separating the kidney from the costodiaphragmatic recess.

FIG. 8–17 [printable version of FIG 8-17]
FIG. 8–18
SECTION 31
A section through the phallus, caudal part of the testis and kidney and the L-1 spinal ganglion.
Observe:
- The corpus cavernosum in the phallus lateral to which is the scrotal swelling at the level of the pubic symphysis.
- The mesonephric duct and regressing female duct in the broad fold.
- The gubernacular fold coursing from the caudal pole of the testis.
- The caudal part of the mesonephros that gives rise to the paradidymis.
- The ganglion cells ventral to the aortic bifurcation.
A section through the glans penis, urethral plate and L-1 and L-2 spinal ganglia.
Observe:
- The corpus spongiosum on each side of the urethral plate.
- The retropubic space between the pubic symphysis and bladder.
- The ureter in two locations, one dorsal to the mesonephric duct, the other at its junction with the bladder lining.
- The femoral nerve between the psoas major and iliacus muscles.
- The junction of the common iliac veins forming the inferior vena cava.

FIG. 8–18 [printable version of FIG 8-18]
FIG. 8–19
SECTION 33
A section through the bulb and crura of the penis and L-2 spinal ganglion.
Observe:
- The proximal part of the urethra where the prostate will later develop.
- The mesonephric ducts in the fused broad folds.
- The caudal edge of the ureter.
- The junction of the colon in Section 32 with the rectum.
- The internal iliac vessels lateral to the pelvic viscera.
A section through the urogenital groove and fold and L-3 spinal ganglion.
Observe:
- The three cartilagenous precursors of the pelvic bone; ilium, ischium and pubis.
- The near junction of the mesonephric ducts with the urethra.
- The fused female ducts in the midline.
- The rectovesicle pouch of the peritoneal cavity.
- The ganglion cells of the pelvic plexus lateral to the pelvic viscera.

FIG. 8–19 [printable version of FIG 8-19]
FIG. 8–20
SECTION 35
A section through the hip joint and L-3 spinal ganglion.
Observe:
- The junction of the urethra with the urogenital groove.
- The caudal part of the peritoneal cavity (rectovesicle pouch).
- The absence of a mesentery on the rectum.
- An intervertebral disc.
- The subdivisions of the white and grey matter in the lumbar portion of the spinal cord.
A section through the anus, anal canal, caudal part of the rectum and L-3 and L-4 spinal ganglia.
Observe:
- The ischiorectal fossa lateral to the rectum and anal canal between the levator ani and obturator internus muscles.
- The sympathetic trunk medial to the S-1 ventral primary ramus.
- The sacral plexus between the rectum and the greater sciatic foramen.
- The ala of the sacrum medial to the sacroiliac joint.
- The gluteal muscles in the buttock.
A section through the coccyx and S-1 and S-2 spinal ganglia.
Observe:
- The termination of the primitive filum terminale dorsal to the coccyx.
- The termination of the sympathetic trunks in Section 36 in the ganglion impar ventral to the coccyx.
- The three layers of the sacral part of the spinal cord.
A section through the terminal ventricle of the central canal.
Observe:
- The two derivatives of the ectomeninx: the chondrogenic layer and dura mater separated by a venous plexus.
- The junction of the primitive filum terminale with the spinal cord.

FIG. 8–20 [printable version of FIG 8-20]