Developmental Stages in Human Embryos
Go to Stage: Intro 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
Preface | Introduction
DEVELOPMENTAL STAGES IN HUMAN EMBRYOS
Including a Revision of Streeter's "Horizons" and a Survey of the Carnegie Collection
RONAN O'RAHILLY
and
FABIOLA MÜLLER
Carnegie Laboratories of Embryology,
California Primate Research Center,
and
Departments of Human Anatomy and Neurology,
University of California, Davis

CARNEGIE INSTITUTION OF WASHINGTON
PUBLICATION 637
1987
Dem Andenken von Wilhelm His, dem Älteren, der vor hundert Jahren die Embryologie des Menschen einführte
und dem seines Protégé, Franklin P. Mall, dem Begründer der Carnegie Collection.

Wilhelm His, 1831-1904

Franklin P. Mall, 1862-1917

George L. Streeter, 1873-1948
PREFACE
During the past one hundred years of human embryology, three landmarks have been published: the Anatomie der menschlichen Embryonen of His (1880–1885), the Manual of Human Embryology by Keibel and Mall (1910–1912), and Streeter's Developmental Horizons in Human Embryos (1942–1957, completed by Heuser and Corner). Now that all three milestone volumes are out of print as well as in need of revision, it seems opportune to issue an updated study of the staged human embryo.
The objectives of this monograph are to provide a reasonably detailed morphological account of the human embryo (i.e., the first eight weeks of development), a formal classification into developmental stages, a catalogue of the preparations in the Carnegie Collection, and a reference guide to important specimens in other laboratories. The Carnegie staging system has now been accepted internationally and, when carefully applied, allows detailed comparisons between the findings at one institution and those at another.
The classifications and descriptions of stages 1–9 are based on light microscopy, and the criteria selected have stood the test of time. The present account is a revision of O'Rahilly's monograph of 1973. In stages 10–23, increasing attention is paid to external form, although internal structure is not, and should not be, neglected. Streeter's masterly account has been updated and is used here in treating stages 10–23. Many of Streeter's paragraphs have been left virtually unchanged (except for improvements in terminology) but others have been altered considerably, and much new material has been added.
Most of the drawings for stages 1–9 were prepared in the Department of Art as Applied to Medicine, the Johns Hopkins School of Medicine, under the direction of Ranice Crosby. Most of those for stages 10–23 are the work of James F. Didusch, although certain modifications in his terminology have been adopted.
A major change made here from Streeter's account is in the systematic inclusion of standard references. It should be stressed, however, that no attempt has been made to provide a comprehensive bibliography. Many more references can be found in a series of articles on the timing and sequence of developmental events, beginning in Acta anatomica in 1971 and continuing in the Zeitschrift für Anatomie und Entwicklungsgeschichte (now Anatomy and Embryology) from 1971 to 1983. For the nervous system, further references can be found in a continuing series of articles that began in 1981 in Anatomy and Embryology: stages 8–11 have already been published.
Particular attention has been paid to nomenclature throughout. Most of the terms used are in agreement with the Nomina embryologica. The latter, however, unlike the Nomina anatomica, is not exclusively human, and hence certain inappropriate terms have been replaced here.
It is appropriate to acknowledge here the great help provided by the National Institutes of Health, which have supported the writers' research (Grant No. HD-16702, Institute of Child Health and Human Development).
It is a particular pleasure to acknowledge the enthusiasm and collaboration of the late Dr. Ernest Gardner over many years, the friendship and assistance provided by Dr. Elizabeth M. Ramsey, and the continued encouragement of Dr. James D. Ebert, President of the Carnegie Institution of Washington, whose invitation to establish the first nine stages was made twenty years ago.
The authors wish to thank Mr. Ray Bowers and Miss Patricia Parratt of the Carnegie Institution's publications office for the exceptionally great care with which they brought this monograph to fruition.
R. O'R.
F. M.
January 1987
Page 1
INTRODUCTION
embryos should be arranged in stages.
FRANKLIN P. MALL
The combined use of fixation, sectioning with a microtome, and reconstruction from the resultant sections first enabled Wilhelm His, Senior, to begin to elucidate thoroughly the anatomy of individual human embryos. Indeed, His may rightfully be called the “Vesalius of human embryology” (Müller and O'Rahilly, 1986a).
Although fixatives other than spirits were introduced early in the nineteenth century, formalin was not employed until the 1890s. His devised a microtome in about 1866. (A microtome had already been employed as early as 1770.) The wax plate reconstruction technique of Born (1883), introduced in 1876, has undergone numerous modifications over the years. These, as well as graphic reconstruction, have been discussed in a number of publications, e.g., by Gaunt and Gaunt (1978). Florian, who used graphic reconstruction of the human embryo to great advantage, elaborated the mathematical background in Czech in 1928. (See also Fetzer and Florian, 1930.)
It has been pointed out that “the idea of working out a complete account of the development of the human body was always before the mind of His,” and his collaborator, Franz Keibel, proposed to provide “an account of the development of the human body, based throughout on human material” (Keibel and Mall, 1910) rather than from the comparative standpoint. The result was the Manual of Human Embryology edited by Keibel and Mall (1910, 1912), which was an important step in the goal of seeking precision in human embryology. The hope was expressed that, subsequently, a second attempt, “whether made by us or by others, will come so much nearer the goal” (ibid.).
The Carnegie Collection
Mall's collection of human embryos, begun in 1887, later became the basis of the Carnegie Collection (Mall and Meyer, 1921). Mall (1913) stated his indebtedness to His in the following terms: “We must thank His for the first attempt to study carefully the anatomy of human embryos, but his work was planned on so large a scale that he never completed it…. Thus we may trace back to him the incentive for Keibel's Normentafeln, Minot's great collection of vertebrate embryos and mine of human embryos.”
In more recent years the Carnegie Collection has benefited enormously from the meticulous investigations of Bartelmez, the technical adroitness of Heuser, and the donation of, as well as research on, remarkably young specimens by Hertig and Rock. The microtomy of Charles H. Miller and William H. Duncan, the reconstructions by Osborne O. Heard, the artwork by James F. Didusch, and the photography of Chester F. Reather and Richard D. Grill, have each played a key role in the establishment of the superb embryological collection on which the present monograph is so largely based. In George W. Corner's apt comparison, the Collection serves “as a kind of Bureau of Standards.”
Embryological Seriation
His had made the first thorough arrangement of human embryos in the form of a series of selected individual embryos, numbered in the presumed order of their development. The same principle was followed in the published plates known as the Normentafeln, Page 2 edited by Franz Keibel from 1897 onward; the volume on the human (by Keibel and Elze) appeared in 1908. The limitations of the method are (l) that individual embryos cannot be arranged in a perfect series, because any given specimen may be advanced in one respect while being retarded in another, and (2) that it may prove impossible to match a new embryo exactly with any one of the illustrated norms. The need for a more flexible procedure than a mere Entwicklungsreihe soon became apparent in experimental embryology.
Embryonic Staging
In the words of Ross G. Harrison (Wilens, 1969), “the need for standardized stages in the embryonic development of various organisms for the purpose of accurate description of normal development and for utilization in experimental work has long been recognized.” Because “development is a continuous process with an indefinite number of stages” (ibid.),a certain number have to be chosen. Thus each stage “is merely an arbitrarily cut section through the time-axis of the life of an organism” (deBeer, 1958). It resembles, in Harrison's apt comparison, a frame taken from a cine-film. Stages are based on the apparent morphological state of development, and hence are not directly dependent on either chronological age or on size. Furthermore, comparison is made of a number of features of each specimen, so that individual differences are rendered less significant and a certain latitude of variation is taken into account.
Although embryonic staging had been introduced toward the end of the nineteenth century, it was first employed in human embryology by Franklin P. Mall (1914), founder of the Department of Embryology of the Carnegie Institution of Washington.
On the basis of photographs of their external form, Mall (1914) arranged 266 human embryos 2–25 mm in length in a series of fourteen stages, lettered from H to U. (A to G were to have been the earlier stages.)
Mall's successor, George L. Streeter, provided the definitive classification of human embryos into stages, which he termed “developmental horizons.” Attention was concentrated on embryos up to about 32 mm greatest length because it was believed that, during the fetal period, the rate of increment in size and weight might be large enough to provide an adequate index of relative development.
The original plan was “to cover as far as possible the earliest specimens up to fetuses between 32 and 38 mm. long, the stage at which the eyelids have come together,” and “twenty-five age groups” were envisioned (Streeter, 1942). Subsequently, Streeter (1951) decided that stage 23 “could be considered to mark the ending of the embryonic period” proper. The onset of marrow formation in the humerus was “arbitrarily adopted as the conclusion of the embryonic and the beginning of the fetal period of prenatal life. It occurs in specimens about 30 mm. in length” (Streeter, 1949). A scheme of the 23 stages, as modified and used in the present monograph, is provided in Table 0-1.
The term “horizon” was borrowed from geology and archaeology by Streeter (1942) in order “to emphasize the importance of thinking of the embryo as a living organism which in its time takes on many guises, always progressing from the smaller and simpler to the larger and more complex.” However, the somewhat infelicitous term “horizon” has now been replaced by “stage” because the latter is the simple term employed for all other vertebrate embryos. Not only was the term “stage” used decades ago by Harrison for Ambystoma and subsequently by Hamburger and Hamilton for the chick embryo, as well as by others for a variety of reptiles, birds, and mammals, but, even in the case of the human, the term “stage” was employed by Mall (1914) when he first staged the human embryo more than half a century ago. The term is simpler, clearer, of widespread usage, and can be employed as a verb (to stage an embryo) as well as a participial adjective (a staging system). Furthermore, it should be pointed out that such expressions as “at the 3-mm stage” should be replaced by “at 3 mm.” In other words, the length of an embryo is a single criterion that is not in itself sufficient to establish a stage. The term “stage” should be confined to its present-day usage in embryology (such as the 46 stages of Hamburger and Hamilton in the chick, and the 46 stages of Harrison in Ambystoma maculatum).
Additional alterations that have been made in the current work include the replacement of Roman by Arabic numerals and the elimination of the scientifically meaningless term “ovum.” Atlases based on the Carnegie system of staging have Page 3 been prepared by Blechschmidt (1973) and by Gasser (1975). Alternative systems of staging (discussed by O'Rahilly, 1973) are now obsolescent.
Stages 10–23 were published either by Streeter (1942, 1945, 1948, and 1951) or at least with the aid of his notes (Heuser and Corner, 1957). “The earliest age groups” were wisely “to be reserved to the last, so that advantage may be taken of any new material that becomes available” (Streeter, 1942). These groups, stages 1–9, which were to have been completed by the late Chester H. Heuser, became the task of O'Rahilly (1973).
Embryonic Length
Because most embryos are received already in fixative, it is more practicable for comparisons to use measurement after fixation as the standard (Streeter, 1945). The most useful single measurement is the greatest length (G.L.) of the embryo as measured in a straight line (i.e., caliper length) without any attempt to straighten the natural curvature of the specimen (Mall, 1907) and preferably (for purposes of standardization) after two weeks in 10 percent formalin (Streeter, 1920). Up to stage 10, measurements are frequently made on accurately scaled models, although the results (because of shrinkage in preparing the sections) are then smaller (by 25 percent, according to Streeter, 1942). A particularly interesting study has been made of the shrinkage of (pig) embryos in the procedures preparatory to sectioning (Patten and Philpott, 1921). Careful technique (see Heard, 1957) is naturally to be encouraged in order to keep artifactual changes to a minimum.
Table 0-1. Developmental Stages in Human Embryos
| Carnegie Stage |
Pairs of Somites |
Size (mm) |
Age* (days) |
Features |
|---|---|---|---|---|
| 1 | 0.1-0.15 | 1 | Fertilization. | |
| 2 | 0.1-0.2 | 1½-3 | From 2 to about 16 cells. | |
| 3 | 0.1-0.2 | 4 | Free blastocyst. | |
| 4 | 0.1-0.2 | 5-6 | Attaching blastocyst. | |
| 5 | 0.1-0.2 | 7-12 | Implanted although previllous. | |
| 5a | 0.1 | 7-8 | Solid trophoblast. | |
| 5b | 0.1 | 9 | Trophoblastic lacunae. | |
| 5c | 0.15-0.2 | 11-12 | Lacunar vascular circle. | |
| 6 | 0.2 | 13 | Chorionic villi; primitive streak may appear. | |
| 6a | Chorionic villi. | |||
| 6b | Primitive streak. | |||
| 7 | 0.4 | 16 | Notochordal process. | |
| 8 | 1.0-1.5 | 18 | Primitive pit; notochordal and neurenteric canals; neural folds may appear. | |
| 9 | 1-3 | 1.5-2.5 | 20 | Somites first appear. |
| 10 | 4-12 | 2-3.5 | 22 | Neural folds begin to fuse; 2 pharyngeal bars; optic sulcus. |
| 11 | 13-20 | 2.5-4.5 | 24 | Rostral neuropore closes; optic vesicle. |
| 12 | 21-29 | 3-5 | 26 | Caudal neuropore closes; 3-4 pharyngeal bars; upper limb buds appearing. |
| 13 | 30-? | 4-6 | 28 | Four limb buds; lens disc; otic vesicle. |
| 14 | 5-7 | 32 | Lens pit and optic cup; endolymphatic appendage distinct. | |
| 15 | 7-9 | 33 | Lens vesicle; nasal pit; antitragus beginning; hand plate; trunk relatively wider; future cerebral hemispheres distinct. | |
| 16 | 8-11 | 37 | Nasal pit faces ventrally; retinal pigment visible in intact embryo; auricular hillocks beginning; foot plate. | |
| 17 | 11-14 | 41 | Head relatively larger; trunk straighten nasofrontal groove distinct; auricular hillocks distinct; finger rays. | |
| 18 | 13-17 | 44 | Body more cuboidal; elbow region and toe rays appearing; eyelid folds may begin; tip of nose distinct; nipples appear; ossification may begin. | |
| 19 | 16-18 | 47½ | Trunk elongating and straightening. | |
| 20 | 18-22 | 50½ | Upper limbs longer and bent at elbows. | |
| 21 | 22-24 | 52 | Fingers longer; hands approach each other, feet likewise. | |
| 22 | 23-28 | 54 | Eyelid and external ear more developed. | |
| 23 | 27-31 | 56½ | Head more rounded; limbs longer and more developed. |
*Olivier and Pineau (1962) for stages 11-23. Miscellaneous sources for stages 1-10.
Page 4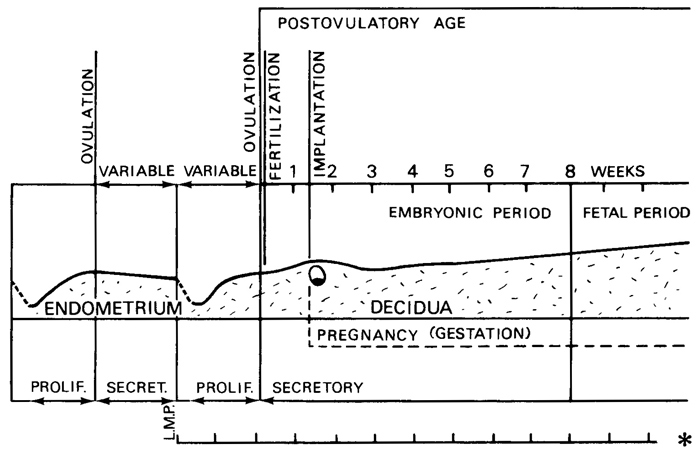
Fig. 0-1. Diagram of endometrial-decidual and embryonic-fetal relationships in relation to time. The second ovulation shown, which is followed by fertilization, is that from which postovulatory age is calculated. The last menstrual period (L.M.P.), which occurred a variable time previously, marks the beginning of the “gestational interval” (asterisk), as defined by Treloar, Behn, and Cowan (1967), who consider pregnancy (gestation) to begin with implantation, whereas others use fertilization as the starting point. Postconceptual hemorrhage in phase with menstruation would result in an apparently short gestational interval. On the other hand, an unrecognized abortion preceding pregnancy, if no menstruation intervened, would result in an apparently long gestational interval. Such possibilities, together with variability in both premenstrual and postmenstrual phases of the cycle, render menstrual data unsatisfactory in the assessment of embryonic age.
The crown-rump (C.-R.) length appears to have been introduced into embryology by Arnold in 1887 (Keibel and Mall, 1910), although the sitting height had been used as a measurement in the adult by Leonardo da Vinci. In the human embryo, from about stage 12 onward it becomes practicable to use the C.-R. length, but this measurement is less satisfactory than, and should be replaced by, the G.L., which can be used from stage 6 throughout the remainder of the embryonic and also the fetal period (O'Rahilly and Müller, 1984a). Other mensural criteria, such as foot length during the fetal period, may be employed, particularly if the specimen has been damaged. The G.L. (like the C.-R. length) should always be stated in millimeters. Particularly in the case of larger embryos and all fetuses, the G.L. of a given specimen should always be stated in preference to, or at least in addition to, its supposed age.
The embryonic lengths given in Table 0-1 indicate the suggested norms. Where possible they are based on specimens graded as excellent and after fixation. It should be stressed, however, that the figures do not indicate the full range within a given stage, especially when specimens of poor quality are included.
Body weight has been somewhat neglected within the embryonic period proper, although some data are available (Witschi, 1956a; Jirásek, Uher, and Uhrová, 1966; Nishimura et al., 1968). By stage 23, the embryo weighs about 2–2.7 grams.
Embryonic Age
The supposed age, as dubiously estimated from the menstrual history, is seldom useful within the embryonic period proper, and such expressions as “at the 18-day stage” should have no place in present-day embryology. Moreover, allowance should be made, but generally is not, for considerable variability in both Page 5 premenstrual and postmenstrual (Stewart, 1952) phases of the menstrual cycle (Vollman, 1977), as well as for the possibility of incorrect identification of menstruation or erroneous interpretation of its absence (Treloar, Behn, and Cowan, 1967).
The ages of very early human embryos (those of the first 3–4 weeks) have been estimated chiefly by comparing their development with that of monkey conceptuses of known postovulatory ages (Rock and Hertig, 1944). Coital history, the condition of the corpus luteum, and the appearance of the endometrium are also taken into account (Rock and Hertig, 1948). More recently, ovulatory tests are providing additional information.
When an embryo has been staged, its presumed age in postovulatory days can be gauged from an appropriate table. The term “postovulatory age” (fig. 0-1) refers to the length of time since the last ovulation before pregnancy began. Because fertilization must occur very close to the time of ovulation, the postovulatory interval is a satisfactory indication of embryonic age. “Menstrual age,” on the other hand, is a misnomer in that it does not indicate age. Furthermore, for precise timing, the words “gestation,” “pregnancy,” and “conception” should be avoided because fertilization is not universally accepted as the commencement: some authors use implantation.
In Table 0-1, the ages are based on Hertig, Rock, and Adams (1956) for stages 2–7, on Heuser (1932a) for stage 8, on Ludwig (1928) for stage 9, on Heuser and Corner (1957) for stage 10, and on Olivier and Pineau (1962) for stages 11–23. The range is not indicated but (at least for stages 10–23) it was believed by Streeter to be ±1 day for any given stage. It should be noted, however, that from stage 14 onward, the ages become increasingly greater than those given by Streeter, which were based on comparisons with macaque embryos; it is now known that such comparisons are not warranted at these stages. Thus, by the time that the embryo reaches stage 23, there is general agreement that it is not 47 ± 1 days (Streeter, 1951) but rather at least 56 days (Witschi, 1956; Olivier and Pineau, 1962; Jirásek, Uher, and Uhrová, 1966; Jirásek, 1971). It has been confirmed ultrasonically in vivo that an embryo of 30 mm is normally aged 8 postovulatory weeks (Drumm and O'Rahilly, 1977).
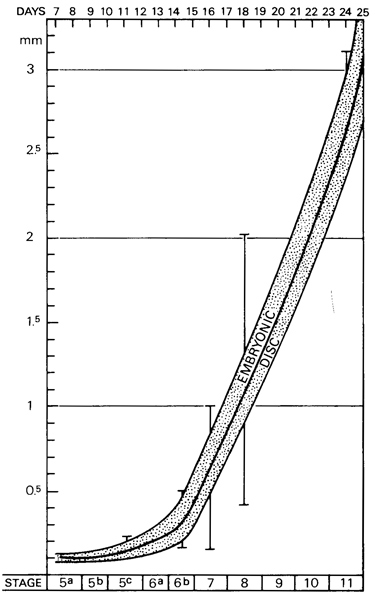
Fig. 0-2. The length of the embryonic disc from stage 5 to stage 11, approximately 1–3½ postovulatory weeks, based on the measurements of 81 embryos. Most of the specimens may be expected to fall within the shaded band, but extreme values are indicated by the five vertical lines. At 1 week the diameter of the disc is approximately 0.1 mm. At 2 weeks the disc is about 0.2 mm in length. At 3 weeks the embryonic length has increased to about 1.5–2 mm. The measurements at later stages are shown in figure 0-3.
Page 6
Fig. 0-3. The length of the embryo from stage 8 to stage 23, approximately 2½to 8 postovulatory weeks, based on the measurements of more than 100 specimens that had been graded as excellent in quality. The measurements at earlier stages are shown in figure 0-2. The maximum diameter of the chorion has also been included (based on 200 specimens graded as either good or excellent): the shaded band includes approximately 80 percent of the specimens. At 4 weeks the embryo is about 5 mm in length and the chorion about 25 mm in diameter. At 8 weeks the embryo is about 30 mm in length, and the chorion is about 65 mm in diameter.
A table of “gestational age . . . estimated from anamnestic data available for embryos in the author's collection” has been published by Jirásek (1971). With few exceptions (chiefly stage 15), the figures given by Jirásek resemble closely those provided by Olivier and Pineau. Although in a few instances (stages 17, 19, 20, and 21) Jirásek’s ages are from one-half to one day older, in most cases the figures of Olivier and Pineau fall within the range listed by Jirásek.
Further details concerning menstrual data and embryonic age have been provided by Moore, Hutchins, and O'Rahilly (1981). Human growth during the embryonic period proper has been discussed by O'Rahilly and Müller (1986a).
Page 7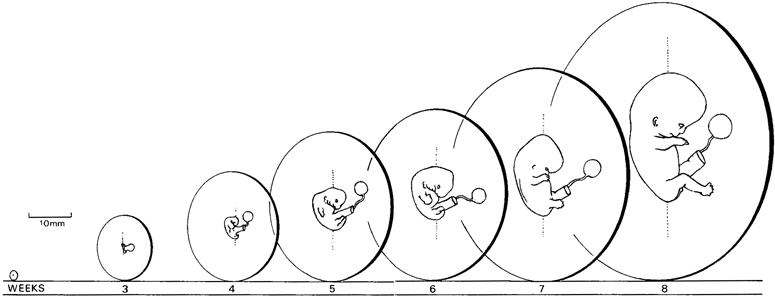
Fig. 0-4. The relative size of the embryo and the chorion at weekly intervals. The stages shown are 6, 10, 13, 16, 17, 20, and 23. The drawings are at approximately the scale of the actual specimens.
Normality
The majority of the approximately 600 sectioned Carnegie embryos assigned to the 23 stages are listed as normal, although variations in, and even anomalies of, individual organs are known to occur. It should not be assumed, however, that every minor defect would necessarily lead to a recognizable anomaly in later life. In the present investigation, an effort has been made to note specifically the presence of frankly abnormal specimens. Nevertheless, it is still true that “as our knowledge of the normal becomes more complete, we find that more and more young embryos which formerly were regarded as normal are not really so ... it remains impossible even at the present time, to determine in all cases whether we are dealing with a normal or an abnormal specimen, even after it has been mounted in serial sections” (Meyer, in Mall and Meyer, 1921). It may be concluded that “The Borderland of Embryology and Pathology” (Willis, 1962) continues to be an important and fruitful area of investigation.
Terminology
In accordance with current practice in anatomical nomenclature, eponyms are avoided wherever possible.
The term anterior and posterior should never be used for the early embryonic period (stages 1–12) and are best avoided until considerably later. Terms considered unsuitable are listed in Table 0-2, together with suggestions for their replacement. Unfortunately the Nomina embryologica contains a number of inappropriate terms.
Table 0-2. List of Discarded and Replaced Terms
| Alternative, Inappropriate, or Incorrect Terms |
Terms Used Here |
|---|---|
| Blastocoel | Blastocystic cavity |
| Blastopore | Primitive pit |
| Blastula | Blastocyst |
| Branchial | Pharyngeal; visceral |
| Chorda dorsalis | Notochord |
| Embryonic shield | Embryonic disc (including cloacal membrane) |
| Formative cells | Epiblast |
| Gastrulation | Not used |
| Germ disc | Epiblast |
| Gestational age | Not used |
| Head process | Notochordal process |
| Horizon | Stage |
| Medullary- groove and folds | Neural groove and folds |
| Menstrual age | Not used |
| Morula | Late stage 2 embryo |
| Ovum (egg) | Oocyte; ootid; embryo |
| Perivitelline space | Subzonal space |
| Placode | Plate or disc |
| Pronephros | Rostralmost part of mesonephros |
| Tail | Not used |
| Ultimobranchial | Telopharyngeal |
| Vitellus | Ooplasm; cytoplasm |
| Yolk sac | Umbilical vesicle |
Page 8
Graphs
Embryonic length is shown in figures 0-2 and 0-3, and the approximate diameter of the chorion is also provided in the latter. Relative sizes at weekly intervals are illustrated in figure 0-4.
Extension of Carnegie System
The attractive idea of using a standard system of staging throughout vertebrate embryology, as espoused by Emil Witschi, has been furthered by the recent appearance of an atlas for staging mammalian and chick embryos, based on the Carnegie system (Butare and Juurlink, 1987).
Copyright © 1987 Carnegie Institution of Washington. Reproduced on ehd.org with permission.